Resumos
A estrutura do solo é constituída de agregados pedogênicos e biogênicos, formados respectivamente pela hierarquização de agregados e pela ação da macrofauna, sobretudo a das minhocas. Neste estudo, objetivou-se a comparação entre agregados do solo (pedogênicos) e coprólitos de minhocas (biogênicos) de diferentes tamanhos e classes de solos. As coletas de amostras indeformadas de solos e coprólitos de minhocas (Pontoscolex corethrurus Muller, 1857) foram realizadas em quatro solos de classes diferentes do Estado da Paraíba (Latossolo Amarelo, Argissolo Vermelho Amarelo, Luvissolo Crômico e Nitossolo Vermelho). As caracterizações físicas e químicas de agregados e coprólitos foram realizadas após secagem à sombra e separação em três classes de diâmetros (20,0 - 9,52; 9,52 - 6,35 e 6,35 - 4,76 mm). A granulometria mostrou ser similar para cada tipo de agregado (pedogênico e biogênico), independentemente do tamanho dos agregados. Em geral, os coprólitos apresentaram maiores proporções de argila, silte e areia fina e menor proporção da fração areia grossa, maior estabilidade e maiores teores de carbono orgânico e cátions trocáveis, comparativamente aos agregados pedogênicos. Os resultados refletem um processo genético peculiar e sugerem a importância dos agregados biogênicos como indicadores da qualidade do solo.
Macrofauna; estabilidade de agregados; atividade biológica; qualidade do solo
The soil structure is composed by pedogenic and biogenic aggregates, formed respectively by aggregate hierarchy and macrofauna activity, especially by earthworms. The objective of this study was to compare soil aggregates and earthworm casts with different aggregate-size classes and soil classes. The sampling of undisturbed soil and earthworm casts (Pontoscolex corethrurus, Muller, 1857) was made in four soils of different classes in the State of Paraíba (Oxisol, Ultisol, Alfisol and Nitisol). The chemical and physical evaluations of aggregates and earthworm casts were determined after dry and split into three aggregate-size classes (20.0 to 9.52, 9.52 to 6.35 and 6.35 to 4.76 mm). The particle-size distribution was similar to aggregates and earthworm casts, independent of aggregate-size classes. Earthworm casts had higher proportions of clay, silt and fine sand, lower coarse sand, higher physical stability and more organic carbon and cations contents than the pedogenic aggregates. These results show a peculiar genetic process that makes biogenic aggregates an important indicator of soil quality.
Macrofauna; aggregate stability; biologic activity; soil quality
CIENCIAS AGRÁRIAS
Luís de França da Silva NetoI; Ivandro de França da SilvaII; Alberto Vasconcellos IndaIII; Paulo César do NascimentoIII; Leandro BortolonIII
IUniversidade Federal do Rio Grande do Sul/UFRGS Departamento de Solos Avenida Bento Gonçalves 9500 91501-970 Porto Alegre, RS luisdefranca@yahoo.com.br
IIUniversidade Federal da Paraíba/UFPB Departamento de Solos e Engenharia Rural Areia, PB
IIIUniversidade Federal do Rio Grande do Sul/UFRGS Departamento de Solos Porto Alegre, RS
RESUMO
A estrutura do solo é constituída de agregados pedogênicos e biogênicos, formados respectivamente pela hierarquização de agregados e pela ação da macrofauna, sobretudo a das minhocas. Neste estudo, objetivou-se a comparação entre agregados do solo (pedogênicos) e coprólitos de minhocas (biogênicos) de diferentes tamanhos e classes de solos. As coletas de amostras indeformadas de solos e coprólitos de minhocas (Pontoscolex corethrurus Muller, 1857) foram realizadas em quatro solos de classes diferentes do Estado da Paraíba (Latossolo Amarelo, Argissolo Vermelho Amarelo, Luvissolo Crômico e Nitossolo Vermelho). As caracterizações físicas e químicas de agregados e coprólitos foram realizadas após secagem à sombra e separação em três classes de diâmetros (20,0 - 9,52; 9,52 - 6,35 e 6,35 - 4,76 mm). A granulometria mostrou ser similar para cada tipo de agregado (pedogênico e biogênico), independentemente do tamanho dos agregados. Em geral, os coprólitos apresentaram maiores proporções de argila, silte e areia fina e menor proporção da fração areia grossa, maior estabilidade e maiores teores de carbono orgânico e cátions trocáveis, comparativamente aos agregados pedogênicos. Os resultados refletem um processo genético peculiar e sugerem a importância dos agregados biogênicos como indicadores da qualidade do solo.
Termos para indexação: Macrofauna, estabilidade de agregados, atividade biológica, qualidade do solo.
ABSTRACT
The soil structure is composed by pedogenic and biogenic aggregates, formed respectively by aggregate hierarchy and macrofauna activity, especially by earthworms. The objective of this study was to compare soil aggregates and earthworm casts with different aggregate-size classes and soil classes. The sampling of undisturbed soil and earthworm casts (Pontoscolex corethrurus, Muller, 1857) was made in four soils of different classes in the State of Paraíba (Oxisol, Ultisol, Alfisol and Nitisol). The chemical and physical evaluations of aggregates and earthworm casts were determined after dry and split into three aggregate-size classes (20.0 to 9.52, 9.52 to 6.35 and 6.35 to 4.76 mm). The particle-size distribution was similar to aggregates and earthworm casts, independent of aggregate-size classes. Earthworm casts had higher proportions of clay, silt and fine sand, lower coarse sand, higher physical stability and more organic carbon and cations contents than the pedogenic aggregates. These results show a peculiar genetic process that makes biogenic aggregates an important indicator of soil quality.
Index terms: Macrofauna, aggregate stability, biologic activity, soil quality.
INTRODUÇÃO
A teoria clássica para a gênese da estrutura do solo tem sido a da hierarquização de agregados (Tisdall & Oades, 1982), segundo a qual os agregados classificados quanto ao tamanho se unem sucessivamente no solo, por meio de diferentes agentes cimentantes originando agregados de maior tamanho e complexidade. Na formação pedogênica dos agregados estão envolvidos fatores do solo como: textura, cátions polivalentes, óxidos e hidróxidos de ferro e óxidos de alumínio, matéria orgânica, exudatos radiculares, compostos derivados do metabolismo microbiano, ação compressiva de raízes e hifas de fungos e ciclos de umedecimento e secagem do solo. A estrutura do solo possui ainda agregados formados por processos genéticos alternativos, como é o caso dos agregados biogênicos, originários da atividade da macrofauna do solo.
Nesse contexto, as minhocas se destacam como os mais importantes representantes da macrofauna do solo com capacidade de alterar as características pedoambientais. Por meio de seus deslocamentos e atividades, as minhocas produzem agregados e bioporos que afetam propriedades físicas como a estabilidade de agregados, a taxa de infiltração de água no solo, bem como alteram a ciclagem de nutrientes e o crescimento de plantas (Scheu, 2003). O processo de ingestão de solo e resíduos orgânicos pelas minhocas leva à formação de coprólitos, que são agregados biogênicos estáveis e resistentes a ciclos de umedecimento e secagem do solo (Oyedele et al., 2006). Além disso, esse tipo de agregado contribui mais eficientemente na proteção física da matéria orgânica do solo, reduzindo sua taxa de decomposição e elevando o potencial de sequestro de carbono pelo solo.
De acordo com Six et al. (2004), essa estabilidade de agregados é determinada pelas seguintes interações: a) ação mecânica de hifas de fungos originados após excreção dos coprólitos; b) inclusão de polissacarídeos microbianos, estabelecendo fortes ligações entre partículas orgânicas e minerais; c) formação de microagregados organo-minerais servindo-se da matéria orgânica recalcitrante; d) cimentação de partículas do solo por cálcio (ponte de cátions), durante a passagem no trato digestivo das minhocas. Schrader & Zhang (1997) afirmam, ainda, que a estabilidade dos coprólitos é dependente do material de origem do solo e da qualidade da matéria orgânica. Com relação aos elementos químicos do solo, os agregados biogênicos apresentam maiores teores de P, K, Na, Ca, Fe, Cu, Mo, Zn, Mn, e valores de pH superiores aos encontrados no solo (Langebach et al., 2002). A magnitude desses efeitos das minhocas no solo é considerável quando se estima que ingiram uma quantidade de solo entre 200 e 400 Mg ha-1 ano-1 (Lavelle et al., 1983). Contudo, pouca atenção tem sido dada à formação biogênica de agregados, talvez por causa da inabilidade na sua identificação, ao tempo de ciclagem e a sua posição na matriz do solo (Velásquez et al., 2007). Estudos sobre agregação biogênica em diferentes classes de solo e tamanhos de agregados ampliam não apenas o conhecimento da dinâmica desses agregados, mas podem qualificá-los como indicadores potenciais da qualidade do solo. Diante dessa relevância ecológica, neste estudo, objetivou-se comparar atributos físicos e químicos de agregados pedogênicos e biogênicos (coprólitos) de diferentes classes de solos do Estado da Paraíba.
MATERIAL E MÉTODOS
Foram amostradas quatro classes de solos submetidos ao cultivo de gramíneas, sendo: um Latossolo Amarelo distrófico sob cana-de-açúcar e um Argissolo Vermelho Amarelo distrófico sob pastagem de capim braquiária, localizados no município de Areia - PB; e um Nitossolo Vermelho Eutrófico sob pastagem de capim braquiária e um Luvissolo Crômico Pálico abrúptico cultivado com cana-de-açúcar, localizados no município de Alagoinha - PB.
Amostras de solo indeformadas foram coletadas na profundidade de 0 a 20 cm, enquanto que os coprólitos de minhoca (Pontoscolex corethrurus, Muller, 1857) foram coletados por catação na superfície dos diferentes solos com base em sua morfologia distinta. A fim de verificar eventuais diferenças de atributos físicos e químicos do solo em relação ao tamanho de agregados pedogênicos e coprólitos, ambos foram separados em três diferentes classes de diâmetro (20,0 - 9,52; 9,52 - 6,35 e 6,35 - 4,76 mm), utilizando as peneiras de 20,0, 9,52, 6,35 e 4,76 mm de diâmetro de malha. Após secagem à sombra, os agregados pedogênicos e os coprólitos das três classes de diâmetro foram submetidas às análises físicas e químicas. A determinação do diâmetro médio ponderado de agregado úmido (DMPAu) foi realizada de acordo com Tisdall & Oades (1982) e Silva & Mielniczuk (1997). Parte dos agregados pedogênicos e dos coprólitos foi destorroada e passada em peneira de 2,0 mm para obtenção da fração terra fina seca ao ar (TFSA) na qual foram determinadas a granulometria, assim como valores de pH e teores de carbono orgânico (CO), fósforo (P), potássio (K), sódio (Na), cálcio (Ca), magnésio (Mg), alumínio (Al) e acidez potencial (H + Al) conforme Empresa Brasileira de Pesquisa Agropecuária - Embrapa (1997). Foram calculadas também a soma de bases (SB), a capacidade de troca de cátions (CTC) e a saturação por bases (V).
O delineamento experimental utilizado foi inteiramente casualizado, com três repetições. Diferenças estatísticas foram observadas para o grau de dispersão de argila e para os atributos químicos através do teste t (Student) a 5% de probabilidade. A análise das relações entre atributos foi realizada por meio de regressões lineares com auxílio do programa SigmaStat 3.5.
Resultados e discussão
A distribuição das frações granulométricas contidas nos diferentes tamanhos de agregados e coprólitos das quatro classes de solos é apresentada na Figura 1. Nos quatro solos, a granulometria mostrou ser similar para cada tipo de agregado (pedogênico e biogênico), independentemente do tamanho dos agregados. Por outro lado, exceto no Nitossolo Vermelho, foram verificadas diferenças pouco expressivas comparando-se a distribuição granulométrica entre agregados e coprólitos em cada classe de solo. Os coprólitos apresentaram maiores percentuais de silte e areia fina no Latossolo, argila e silte no Argissolo, e silte no Luvissolo; e um menor percentual da fração areia grossa (Figura 1), confirmando resultados obtidos por Oyedele et al. (2006) em condições semelhantes a do presente estudo.
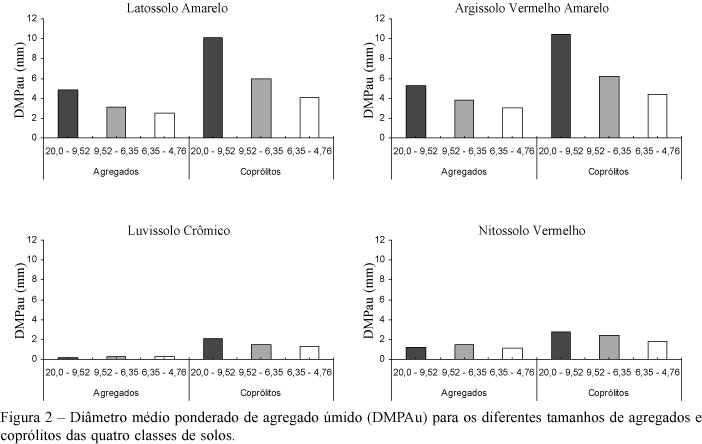
O diâmetro médio ponderado dos agregados por peneiramento úmido (DMPAu) nos diferentes tamanhos de agregados (pedogênicos e biogênicos) distinguiu dois grupos de solos semelhantes (Figura 2), indicando a influência geográfica na formação dos solos nas duas regiões consideradas. No Latossolo e no Argissolo, localizados no município de Areia-PB, os valores de DMPAu foram maiores em relação ao Luvissolo e ao Nitossolo, localizados no município de Alagoinha-PB. Esses resultados estão relacionados com as maiores proporções de argila e as menores proporções de silte e areia muito fina no Latossolo e no Argissolo em comparação ao Luvissolo e ao Nitossolo (Figura 1), conforme indicaram as regressões lineares mostradas no Quadro 1.
Em geral, as frações granulométricas mostraram relações significativas com o DMPAu (Quadro 1), porém com maior grau de significância nos agregados pedogênicos. O teor de CO apresentou relação positiva somente com o DMPAu dos agregados pedogênicos, confirmando observações de Marinissen et al. (1996), que não encontraram relação entre estes atributos em coprólitos. Nestes últimos, é possível que outras variáveis ou interações entre elas atuem de maneira mais expressiva para a estabilização dos agregados. Nos quatro solos, o DMPAu dos coprólitos foi maior que o DMPAu dos agregados (Figura 2), indicando maior estabilidade dos coprólitos quando submetidos à peneiragem por via úmida. Resultados semelhantes foram obtidos por Blanchart et al. (1997), quando compararam agregados de um mesmo solo na presença e ausência de minhocas.
As maiores proporções de argila, cátions e matéria orgânica nos coprólitos podem favorecer interações mais intensas, comparativamente aos agregados pedogênicos. Fatores como a compressão física do solo durante a digestão pelas minhocas podem intensificar os efeitos de polissacarídeos e cátions nas interações organo-minerais (Degens, 1997), especialmente nos coprólitos de classes de tamanho maior. Segundo Oyedele et al. (2006), ao serem ingeridas, as partículas minerais e orgânicas são misturadas com mucos no interior do trato digestivo das minhocas. O processo é acompanhado pela ruptura de ligações minerais (argila-argila) e pela formação de interações (argila-cátion polivalente-matéria orgânica). Esse processo, embora aumente a estabilidade dos coprólitos pode também aumentar o grau de dispersão de argila.
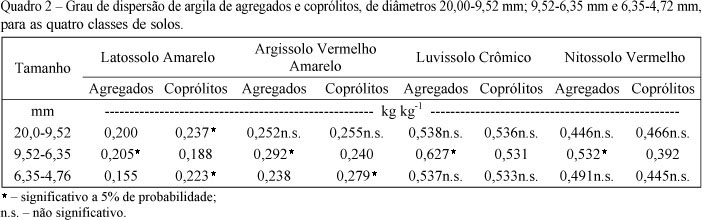
Os valores do grau de dispersão de argila (GD) dos agregados (pedogênicos e biogênicos) para os quatro solos são apresentados no Quadro 2. Na classe de diâmetro entre 9,52 e 6,35 mm dos quatro solos, os agregados pedogênicos apresentaram um maior GD em relação aos coprólitos. Nas demais classes de tamanho avaliadas, houve diferença significativa somente no Latossolo (20,0 9,52 e 6,35 4,76 mm) e no Argissolo (6,35 4,76 mm), com os coprólitos apresentando um maior GD. Considerando o conjunto dos agregados (pedogênicos e biogênicos) dos quatro solos, o GD dos coprólitos foi significativamente menor, em parte, por causa da maior proporção de argila e CO nestes agregados, os quais apresentaram relações negativas com o GD (Quadro 1).
No Quadro 3, são mostrados os valores médios dos atributos químicos dos agregados pedogênicos e coprólitos nas quatro classes de solos. Os coprólitos apresentaram diferenças químicas significativas comparativamente aos agregados pedogênicos. Porém, essas diferenças independem dos tamanhos de agregados e coprólitos considerados neste estudo.
Os valores médios de pH dos coprólitos foram superiores aos dos agregados pedogênicos, fato associado aos menores teores de H+Al e à maior soma de bases nos coprólitos na maioria dos solos. Com relação à bioquímica, essa alteração do pH dos coprólitos pode ser consequência da excreção de amônia no intestino ou da excreção de carbonato de cálcio das glândulas calcíferas na faringe das minhocas, quando o solo é ingerido (Edwards & Bohlen, 1996; Schrader & Zhang, 1997). Os teores médios de Ca dos coprólitos foram superiores aos dos agregados do solo, com destaque para o Argissolo. De acordo com Schrader & Zhang (1997), os coprólitos são enriquecidos pelas glândulas calcíferas durante a passagem no trato digestivo das minhocas. Os teores de Mg foram superiores nos coprólitos, com exceção do Nitossolo, no qual não houve diferença significativa com os teores observados nos agregados pedogênicos.
Os teores de K foram, em média, 5,0 vezes superiores nos coprólitos do Latossolo em comparação aos teores encontrados nos agregados pedogênicos. Teores superiores de K em coprólitos podem ser decorrentes da liberação de K por materiais orgânicos, pela fragmentação ocorrida durante a digestão (Noguera et al., 2010). O Latossolo sob cana-de-açúcar é adubado com NPK anualmente, o que pode ter contribuído para elevar os teores de K nos coprólitos desse solo em comparação com os demais. Consequentemente, a saturação por bases (V) dos coprólitos apresentou valores médios superiores na maioria dos solos, com valores de 42,2% para o Argissolo e 70,6% para o Luvissolo, valores estes 18,8 e 15,5% superiores ao V dos agregados pedogênicos. A dinâmica desses elementos nos coprólitos difere em relação aos agregados pedogênicos não somente pelos teores mais elevados, mas também em virtude do pH mais elevado que afeta a disponibilidade desses elementos (Noguera et al., 2010).
Os teores de P foram superiores nos coprólitos dos quatro solos, com destaque para os teores de P verificados nos coprólitos de Luvissolo, em média 5,0 vezes maiores que os teores de P do solo. Haynes et al. (2003) afirmam que as minhocas ingerem preferencialmente materiais orgânicos ricos em nutrientes, e que o trato digestivo das minhocas é um ambiente que favorece processos bioquímicos e a mineralização por parte dos microrganismos. No caso do Luvissolo, cujos teores de argila são baixos, os valores mais elevados de P nos coprólitos parecem não ter relação com a preferência por materiais orgânicos ricos neste nutriente e sim podem estar relacionados à atividade ácida e básica da fosfatase no intestino das minhocas (Lopez-Hernández et al., 1993). Da mesma forma que o P e o Ca, os teores de CO também foram superiores nos coprólitos de todos os solos. Em termos quantitativos, a diferença média dos teores de CO entre coprólitos e agregados pedogênicos foi superior no Argissolo (6,46 g kg-1) comparativamente aos outros solos, como por exemplo, o Luvissolo (4,12 g kg-1), no entanto, em termos percentuais a diferença entre os teores de CO de agregados e coprólitos foi superior no Luvissolo, uma vez que esse solo apresenta um teor de CO naturalmente baixo (Quadro 3).
Conceitualmente, os coprólitos de minhocas são formados a partir de um processo genético peculiar que determina diferenças nos atributos físicos e químicos comparativamente aos agregados pedogênicos. No presente trabalho, o diâmetro de coprólitos não influenciou nos atributos avaliados, o que os diferencia ainda mais dos agregados pedogênicos formados a partir da hierarquização de agregados. Porém, estudos compartimentalizados, que relacionem a qualidade da matéria orgânica do solo e aspectos mineralógicos, devem ser realizados como propostas futuras. De qualquer maneira, a caracterização física e química de agregados pedogênicos e coprólitos de minhoca permitem inferir sobre a dinâmica diferenciada desses tipos de agregados. Por causa da relevância ecológica da atividade das minhocas no solo e a sua capacidade de alterar o pedoambiente, os agregados biogênicos podem ser considerados como indicadores potenciais da qualidade do solo.
CONCLUSÕES
A distribuição granulométrica indicou que os coprólitos apresentaram maiores proporções de argila, silte e areia fina em comparação com os agregados pedogênicos, indicando a seletividade de partículas como característica pedogenética.
Uma maior estabilidade física dos coprólitos em comparação com os agregados pedogênicos foi indicada pelo maior diâmetro médio de agregados por peneiramento úmido e pelo menor grau de dispersão de argila.
Os coprólitos possuem melhor qualidade dos atributos químicos, representada por maiores valores de pH e teores de cátions, fósforo, carbono orgânico, e menores teores de alumínio e acidez trocável em comparação aos agregados pedogênicos, como um reflexo da atividade biológica na gênese dos agregados biogênicos.
(Recebido em 20 de outubro de 2008 e aprovado em 25 de novembro de 2009)
- BLANCHART, E.; LAVELLE, P.; BRAUDEAU, E.; LE BISSONAIS, Y.; VALENTIN, C. Regulation of soil structure by geophagus earthworm activities in humid savannas of Côte d'Ivoire. Soil Biology Biochemistry, v.29, p.431-439, 1997.
- DEGENS, B.P. Macro-aggregation of soils by biological bonding and binding mechanisms and the factors affecting these: a review. Australian Journal Soil Research, v.35, p.431-459, 1997.
- EDWARDS, C.A.; BOHLEN, P.J. Biology and ecology of earthworms 3.ed. London: Chapman & Hall, 1996.
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA. Centro Nacional de Pesquisa de Solos. Manual de métodos de análise de solo Rio de Janeiro, 1997. 212p. (EMBRAPA-CNPS, Documento, 1).
- HAYNES, R.J.; FRASER, P.M.; PIERCY, J.E.; TREGURTHA, R.J. Casts of Aporrectodea caliginosa (Savigny) and Lumbricus rubellus (Hoffmeister) differ in microbial activity, nutrient availability and aggregate stability. Pedobiologia, v.47, p.882-887, 2003.
- LAVELLE, P.; RANGEL, P.; KANYONYO, J. Mucus production by two species of tropical earthworms: Millsonia lamtoiana (Megascolecidae) and Pontoscolex corethrurus (Glossoscolecidae). In: LEBRUM, P. et al. (Eds.). New trends in soil biology Louvain-la-Neuve: Dieu-Brichard, 1983. p.405-410.
- LOPEZ-HERNÁNDEZ, D.; LAVELLE, P.; FARDEAU, J.C.; NIÑO, M. Phosphorus transformations in two P-sorption contrasting tropical soils during transit through Pontoscolex corethrurus (Glossoscolecidae, Oligichaeta). Soil biology and Biochemistry, v.25, p.789-792, 1993.
- MARINISSEN, J.C.Y.; NIJHUIS, E.; BREEMEN, N. van. Clay dispersability in moist earthworm casts of different soils. Applied Soil Ecology, v.4, p.83-92, 1996.
- NOGUERA, D.; RONDÓNC, M.; LAOSSID, K.R.; HOYOSB, V.; LAVELLEA, P.; CARVALHO, M.H.C.; BAROT, S. Contrasted effect of biochar and earthworms on rice growth and resource allocation in different soils. Soil biology and Biochemistry, v.42, p.1017-1027, 2010.
- OYEDELE, D.J.; SCHJONNING, P.; AMUSAN, A.A. Physicochemical properties of earthworm casts and undingested parent soil from selected sites in southwestern Nigeria. Ecological Engineering, v.28, p.106-113, 2006.
- SCHEU, S. Effects of earthworms on plant growth: patterns and perspectives. Pedobiologia, v.47, p.846-856, 2003.
- SCHRADER, S.; ZHANG, H. Earthworm casting: stabilization or destabilization. Soil Biochemistry, v.29, p.469-475, 1997.
- SILVA, I.F.; MIELNICZUK, J. Ação do sistema radicular de plantas na formação e estabilização de agregados do solo. Revista Brasileira de Ciência do Solo, Viçosa, v.20, n.1, p.113-117, 1997.
- SIX, J.; BOSSUYT, H.; DEGRYZE, S.; DENEF, K. A history on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Research, v.79, p.731, 2004.
- TISDALL, J.M.; OADES, J.M. Organic matter and water-stable aggregates in soils. Journal of Soil Science, v.33, p.141-163, 1982.
- VELÁSQUEZ et al. This ped is my ped: Visual separation and near infrared spectra allow determination of the origins of soil macroaggregates. Pedobiologia, v.51, p.75-87, 2007.
Atributos físicos e químicos de agregados pedogênicos e de coprólitos de minhocas em diferentes classes de solos da Paraíba
Datas de Publicação
-
Publicação nesta coleção
13 Jan 2011 -
Data do Fascículo
Dez 2010
Histórico
-
Recebido
20 Out 2008 -
Aceito
25 Nov 2009