Abstract
The Caatinga is the largest tropical dry forest region in South America and harbors an impressive biological diversity. However, efforts in the studies of many groups of organisms, especially aquatic ones, are very small compared to other Brazilian regions. The state of Ceará stands out due to the major concentration of surface water in the semi-arid region, and the diatom flora is virtually unknown. We performed a pioneering and extensive taxonomic study on the centric diatoms from the Caatinga, documenting the morphological variation of the species found, expanding their geographic distribution, and reviewing taxonomic and nomenclatural aspects when relevant. The study was based on planktonic and periphytic samples collected in four hydrographic basins located in the northwestern region of Ceará, northeastern Brazil. Fifteen infrageneric diatom taxa were identified, described, and illustrated using light and scanning electron microscopy. Discostella stelligera was first recorded for northeastern Brazil and D. woltereckii is a new record to the country. Diminutive centric diatoms are well represented in the samples and their taxonomy is discussed. Terpsinoë musica occurred in a population with high phenotypic plasticity and we discussed the related taxonomic implications.
Keywords:
Bacillariophyta; intermittent water bodies; semiarid; taxonomy
Resumo
A Caatinga é a maior região de floresta tropicalmente seca da América do Sul e abriga uma diversidade biológica impressionante. Porém, os esforços no estudo de diversos grupos de organismos, principalmente aquáticos, são muito escassos se comparados a outras regiões brasileiras. O estado do Ceará destaca-se pela grande concentração de águas superficiais no semiárido e a flora de diatomáceas é praticamente desconhecida. Realizamos um estudo taxonômico pioneiro e extenso sobre as diatomáceas cêntricas da Caatinga, documentando a variação morfológica das espécies encontradas, ampliando sua distribuição geográfica e revisando aspectos taxonômicos e nomenclaturais quando relevantes. O estudo baseou-se em amostras planctônicas e perifíticas coletadas em quatro bacias hidrográficas localizadas na região noroeste do Ceará, nordeste do Brasil. Quinze táxons de diatomáceas infra-genéricos foram identificados, descritos e ilustrados usando microscopia óptica e eletrônica de varredura. Discostella stelligera foi registrada pela primeira vez para o nordeste do Brasil e D. woltereckii é um novo registro para o país. Diatomáceas cêntricas diminutas estão bem representadas nas amostras e sua taxonomia é discutida. Terpsinoë musica ocorreu em uma população com alta plasticidade fenotípica e se discutiram as implicações taxonômicas relacionadas. Palavras-chave: Bacillariophyta, corpos d’água intermitentes, semiárido, taxonomia.
Palavras-chave:
Bacillariophyta; corpos d’água intermitentes; semiárido; taxonomia
Introduction
The Caatinga is an important Brazilian ecological region, recognized as the largest seasonally dry tropical forest in South America ( Silva et al. 2017 Silva JMC, Barbosa LCF, Leal IR & Tabarelli M (2017) The Caatinga: understanding the challenges. In: Silva JMC, Leal IR & Tabarell M (eds.) Caatinga: The largest tropical dry forest region in South America. Springer International Publishing AG, Cham. Pp. 3-19.). It is predominant in the semi-arid region of northeastern Brazil, but is interspersed with remnants of humid and sub-humid areas due to elevation and ocean proximity ( Bastos & Cordeiro 2012Bastos FH & Cordeiro AMN (2012) Fatores naturais na evolução das paisagens no semiárido brasileiro: Uma abordagem geral. Revista Geonorte 3: 464-476.). Studies have demonstrated the vast biodiversity of the Caatinga ( Leal et al. 2005 Leal IR, Silva JMC, Tabarelli M & Lacher T (2005) Changing the course of biodiversity conservation in the Caatinga of Northeastern Brazil. Conservation Biology 19: 701-706.; Albuquerque et al. 2012 Albuquerque UP, Araújo EL, El-Deir ACA, Lima ALA, Souto A, Bezerra BM, Ferraz EMN, Freire EMX, Sampaio EVSB, Las-Casas FMG, Moura GJB, Pereira GA, Melo JG, Ramos MA, Rodal MJN, Schiel N, Lyra-Neves RM, Alves RRN, Azevedo-Júnior SM, Telino-Júnior WR & Severi W (2012) Caatinga revisited: ecology and conservation of an important Seasonal dry Forest. The Scientific World Journal 2012: 1-18.). The fauna and flora are well adapted to water shortages ( Santos et al. 2011 Santos JC, Leal IR, Almeida-Cortez JS, Fernandes GW & Tabarelli M (2011) Caatinga: the scientific negligence experienced by a dry tropical forest. Tropical Conservation Science 4: 276-286.; Silva et al. 2017 Silva JMC, Barbosa LCF, Leal IR & Tabarelli M (2017) The Caatinga: understanding the challenges. In: Silva JMC, Leal IR & Tabarell M (eds.) Caatinga: The largest tropical dry forest region in South America. Springer International Publishing AG, Cham. Pp. 3-19.), but significantly affected by extensive processes of environmental change and deterioration caused by unsustainable use of its resources ( Leal et al. 2003 Leal IR, Silva JMC & Barros MLB (2003) Ecologia e conservação da caatinga. Editora Universitária da UFPE, Recife. 822p.).
The state of Ceará includes a major concentration of surface water in the Caatinga region ( Andrade et al. 2017 Andrade EM, Aquino DN, Chaves LCG & Lopes FB (2017) Water as capital and its uses in the caatinga. In: Silva JMC, Leal IR & Tabarelli M (eds.) Caatinga: the largest tropical dry forest region in South America. Springer International Publishing AG, New York. Pp. 281-302.). A large part of the state has a tropical, semi-arid climate, with high annual average temperatures, high rates of evaporation, and scarce and irregular rainfall that is restricted to a short period of the year ( IPECE 2016IPECE (2016) Características geográficas, recursos naturais e meio ambiente. Ceará em números. Instituto de Pesquisa e Estratégia Econômica do Ceará. Pp. 21-44. Available at < http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2016/completa/>. Access on 25 November 2019.
http://www2.ipece.ce.gov.br/publicacoes/...
). Consequently, most water bodies are seasonally intermittent and many were made permanent by dam construction ( IPECE 2016IPECE (2016) Características geográficas, recursos naturais e meio ambiente. Ceará em números. Instituto de Pesquisa e Estratégia Econômica do Ceará. Pp. 21-44. Available at < http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2016/completa/>. Access on 25 November 2019.
http://www2.ipece.ce.gov.br/publicacoes/...
). Concerning factors are pollution from industry, lack of basic sanitation, siltation resulting from riparian deforestation, and the numerous dams built for water retention ( Moro et al. 2015 Moro MF, Macedo MB, Moura-Fé MM, Castro ASF & Costa RC (2015) Vegetation, phytoecological regions and landscape diversity in Ceará state, northeastern Brazil. Rodriguésia 66: 717-743.). Freshwater sources are threatened by anthropogenic pressures and biodiversity is declining ( Virta et al. 2019 Virta L, Gammal J, Järnström M, Bernard G, Soininen J, Norkko J & Norkko A (2019) The diversity of benthic diatoms affects ecosystem productivity in heterogeneous coastal environments. Ecology 100: 1-11.).
Diatoms are ordinary and abundant components in freshwater systems. They play important ecological roles in aquatic environments as primary producers at the base of the trophic web, which can occur in different nutrient regimes, pH, salinity, and temperature ( Julius & Theriot 2010Julius ML & Theriot EC (2010) The diatoms: a primer. In: Smol JP & Stoermer EF (eds.) The diatoms: applications for the environmental and earth sciences. 2nd ed. Cambridge University Press, Cambridge. Pp. 8-22.). The rapid response time to environmental changes, make diatoms reliable aquatic bioindicators often employed to assess water quality ( Kelly et al.1998 Kelly MG, Cazaubon A, Coring E, Dell’uomo A, Ector L, Goldsmith B, Guasch H, Hürlimann J, Jarlman A, Kawecka B, Kwandrans J, Laugaste R, Lindstrøm EA, Leitao M, Marvan P, Padisák J, Pipp E, Prygiel J, Rott E, Sabater S, Van-Dam H & Vizinet J (1998) Recommendations for the routine sampling of diatoms for water quality assessments in Europe. Journal of Applied Phycology 10: 215-224.; Lobo et al. 2010 Lobo EA, Wetzel CE, Ector L, Katoh K, Blanco S & Mayama S (2010) Response of epilithic diatom communities to environmental gradients in subtropical temperate Brazilian rivers. Limnetica 29: 10-20., 2015Lobo EA, Schuch M, Heinrich CG, Costa AB, Düpont A, Wetzel CE & Ector L (2015) Development of the Trophic Water Quality Index (TWQI) for subtropical temperate Brazilian lotic systems. Environmental Monitoring and Assessment 187: 1-13.; Álvarez-Blanco et al. 2013 Álvarez-Blanco I, Blanco S, Cejudo-Figueiras C & Bécares E (2013) The Duero Diatom Index (DDI) for river water quality assessment in NW Spain: design and validation. Environmental Monitoring and Assessment 185: 969-981.; Lai et al. 2014 Lai GG, Padedda BM, Virdis T, Sechi N & Lugliè A (2014) Benthic diatoms as indicators of biological quality and physical disturbance in Mediterranean watercourses: a case study of the Rio Mannu di Porto Torres basin, northwestern Sardinia, Italy. Diatom Research 29: 11-26.; Stevenson 2014Stevenson J (2014) Ecological assessments with algae: a review and synthesis. Journal Phycology 50: 437-461.; Dalu & Froneman 2016Dalu T & Froneman PW (2016) Diatom-based water quality monitoring in southern Africa: challenges and future prospects. Water SA 42: 551-559.).
In Brazil, diatom research is mostly concentrated in the South and Southeast, due to the greater representation of specialized research centers in these regions ( Menezes et al. 2015 Menezes M, Bicudo CEM & Moura CWN (2015) Update of the Brazilian floristic list of algae and cyanobacteria. Rodriguésia 66: 1047-1062.). Focusing on inventories describing the diversity of diatoms in poorly studied regions of the country is crucial. The earliest studies on diatoms in northeastern Brazil were carried out by Patrick ( 1940aPatrick R (1940a) Diatoms of Northeastern Brazil. Part I. Coscinodiscaceae, Fragilariaceae and Eunotiaceae. Proceedings of the Academy of Natural Sciences of Philadelphia 92: 191-226., bPatrick R (1940b) Some new diatoms from Brazil. Notulae Naturae of the Academy of Natural Sciences of Philadelphia 59: 1-7.), who recorded 30 taxa in Ceará. More recently, only a few studies involving microalgae have been conducted in the southern Ceará, and diatoms have generally been identified to class or genus level, rarely to species level ( Aquino et al. 2011Aquino EP, Oliveira ECC, Fernandes UL & Lacerda SR (2011) Phytoplankton in stabilization ponds in northeast Brazil. Brazilian Journal of Aquatic Sciences and Technology 15: 71-77.; Vieira et al. 2013Vieira RDS, Lacerda SR, Oliveira ECC, Nascimento KJ & Dias AS (2013) Biodiversidade de microalgas perifíticas do Rio da Batateira (Sítio Fundão, Crato, CE). Cadernos de Cultura e Ciência 12: 7-15.; Amorim et al. 2015Amorim CA, Dias AS & Ferreira RJ (2015) Microalgas perifíticas associadas à Nymphoides indica (L.) O. Kuntze em um reservatório do semiárido cearense. Caderno de Cultura e Ciência 14: 43-54.; Costa et al. 2015Costa ARS, Amorim CA, Nascimento KJ, Dias AS, Ferreira RJ & Lacerda SR (2015) Caracterization of microalgae periphytic community in a reservoir semiarid cearense. Caderno de Cultura e Ciência 14: 43-59.). There are approximately 50 taxa of centric diatoms registered in freshwaters from Brazil ( Eskinazi-Leça et al. 2015Eskinazi-Leça E, Moura CWN, Cunha MGGS, Santiago MF, Borges GCP, Lima JC, Silva MH, Ferreira LC, Aquino E, Da Silva WJ & Menezes M (2015) Bacillariophyceae. In: Lista de espécies da flora do Brasil. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Available at < http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB119495>. Access on 28 November 2019.
http://floradobrasil.jbrj.gov.br/jabot/f...
; Ludwig & Tremarin 2017Ludwig TAV & Tremarin PI (2017) Bacillariophyta. In: Bicudo CEM & Menezes M (eds.) Gênero de algas de águas continentais do Brasil: chave para identificação e descrição. 3ª ed. RiMa, Rio de Janeiro. Pp. 435-503.) but there are few records of the centric diatom in the Caatinga region (e.g., Patrick et al. 1940aPatrick R (1940a) Diatoms of Northeastern Brazil. Part I. Coscinodiscaceae, Fragilariaceae and Eunotiaceae. Proceedings of the Academy of Natural Sciences of Philadelphia 92: 191-226., bPatrick R (1940b) Some new diatoms from Brazil. Notulae Naturae of the Academy of Natural Sciences of Philadelphia 59: 1-7.; Tremarin et al. 2013Tremarin PI, Ludwig TV & Torgan LC (2013) Morphological variation and distribution of the freshwater diatom Aulacoseira ambigua (Grunow) Simonsen in Brazilian continental environments. Iheringia Série Botânica 68: 139-157.), highlighting actual regional biodiversity gap and the great need taxonomic studies in that area.
This study aims to expand the knowledge of diatom biodiversity in the state of Ceará reducing gaps in our understanding of the geographic distribution of centric diatoms in the semi-arid region of Brazil. We performed a taxonomic study including samples from four hydrographic basins in northwest Ceará. Herein, we focus on population analysis of the species, detailed descriptions, taxonomic comments, and illustrations using light and electron microscopy.
Material and Methods
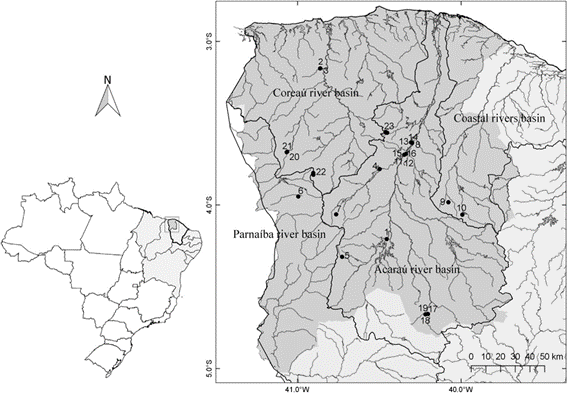
Ceará state is divided into seven mesoregions. The study area is located in the northwest of the state ( Fig. 1) and covers 34,560 km² with a population of 326,847 inhabitants distributed across 47 cities ( IPECE 2008IPECE (2008) características geográficas, recursos naturais e meio ambiente. Ceará em números. Instituto de Pesquisa e Estratégia Econômica do Ceará. Pp. 30-31. Available at < http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2008/completa/>. Access on 28 November 2019.
http://www2.ipece.ce.gov.br/publicacoes/...
), 12 of which were included in this study (Tab. S1, available on supplementary material < https://doi.org/10.6084/m9.figshare.21339363.v1>). Four hydrographic basins make up this mesoregion: Acaraú River, Coreaú River, Coastal rivers, and Parnaíba River ( Nascimento 2011Nascimento F (2011) Categorização de usos múltiplos dos recursos hídricos e problemas ambientais. Revista da Associação Nacional de Pós-graduação e Pesquisa em Geografia 7: 81-97.).

Map of study area: hydrographic basins of northwestern region (in grey) of the state of Ceará, Brazil. Sampling locations (black circles) were numbered from 1 to 23. Information about the sampling points is shown in Table S1, available on supplementary material < https://doi.org/10.6084/m9.figshare.21339363.v1>.
Sampling was conducted during 2016, 2018, and 2019 in different freshwater environments in the river basins (Tab. S1, available on supplementary material < https://doi.org/10.6084/m9.figshare.21339363.v1>). Twenty-three samples were collected: 17 epiphytic, 3 epilithic, 1 epipsamic, and 2 phytoplanktonic. Plankton was obtained with a plankton net (20 µm mesh) and periphyton, which are attached to natural substrates, were collected manually. We measured conductivity and pH, when possible (Tab. S1, available on supplementary material < https://doi.org/10.6084/m9.figshare.21339363.v1>), using conductivity meter CG1400 and pH-meter PG1400, respectively (Gehaka, Ltda).
Samples were preserved in 4% formaldehyde and cleaned according to Simonsen ( 1974Simonsen R (1974) The diatom plankton of the Indian Ocean Expedition of R/V “Meteor”, 1964-65. “Meteor” Forschungsergbnisse, Reih D-Biol 19: 1-66.), with modifications described by Moreira-Filho & Valente-Moreira ( 1981Moreira-Filho H & Valente-Moreira IM (1981) Avaliação taxonômica e ecológica das diatomáceas (Bacillariophyceae) epífitas em algas pluricelulares obtidas nos litorais dos estados dos estados do Paraná, Santa Catarina e São Paulo. Boletim Museu Botânico Municipal de Curitiba 47: 1-17.). Permanent slides were mounted with Naphrax® resin (refractive index = 1.74). Diatoms were analyzed and illustrated using an Olympus BX40 light microscope (Olympus, Tokyo, Japan) equipped with an Olympus DP71 image capture camera. Cleaned sub-samples were deposited in aluminum stubs, coated with gold using a Bal-Tec SCD050 Sputter Coater (Bal-Tec, Balzers, Liechtenstein). Preparations were analyzed in the JEOL JSM 6360-LV (Jeol, Japan) and TESCAN VEGA 3 LMU scanning electron microscopes (Tescan Analytics, Brno, Czech Republic), housed at the Electron Microscopy Center of the Federal University of Paraná (UFPR).
Classification follows Medlin & Kaczmarska ( 2004Medlin LK & Kaczmarska I (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Journal Phycologia 43: 245-270.) for supra-ordinal taxa and Round et al. ( 1990Round FE, Crawford RM & Mann DG (1990) The diatoms: biology and morphology of the genera. Cambridge University Press, Cambridge. 747p.) for subordinal ones. Striae density and marginal processes of centric diatoms were calculated according to Hasle ( 1983Hasle GR (1983) The marine planktonic diatoms Thalassiosira oceanica sp. nov. and T. partheneia. Journal of Phycology 19: 220-229.) and modified by Syvertsen & Hasle ( 1984Syvertsen EE & Hasle GR (1984) Thalassiosira bulbosa Syvertsen sp. nov., an Artic marine diatom. Polar Biology 3: 167-172.). Morphological terminology was based on Round et al. ( 1990Round FE, Crawford RM & Mann DG (1990) The diatoms: biology and morphology of the genera. Cambridge University Press, Cambridge. 747p.), Houk ( 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.) and Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.). Samples and slides were registered and deposited in the herbaria of the Federal University of Paraná (UPCB), Curitiba, Paraná, and the State University of Vale do Acaraú (HUVA), Sobral, Ceará, Brazil (Tab. S1, available on supplementary material < https://doi.org/10.6084/m9.figshare.21339363.v1>).
The frequency of occurrence was calculated according to Dajoz ( 2005Dajoz R (2005) Princípios de ecologia. 7ª ed. Artmed, Porto Alegre. 520p.), as follows: constant (C ≥ 50%), common (C ≥ 30% or ≤ 50%), sporadic (C ≥ 10% or ≤ 30%) and rare (C ≤ 10%) .
Results and Discussion
We present 14 species and one non-typical variety of centric diatoms, distributed among one genera of Coscinodiscophyceae and six genera of Mediophyceae.
Descriptions and comments follow.
Bacillariophyta Karsten Coscinodiscophytina Medlin et Kaczmarska
Coscinodiscophyceae Round et R.M. Crawford Aulacoseirales Crawford
Aulacoseiraceae Thwaites
Aulacoseira Thwaites
Aulacoseira ambigua (Grunow) Simonsen, Bacill. 2:56. 1979Simonsen R (1979) The diatom system: ideas on phylogeny. Bacillaria 2: 9-71.. Fig. 2a-b
Frustules cylindrical; valve face flat with short rows of rounded areolae restricted to the margin ( Fig. 2a); mantle ornamented with striae arranged obliquely in relation to the pervalvar axis in a dextrorse pattern; areolae rounded, delicate. Diameter 5.7–6.4 µm; mantle height 9.9 µm; 17 striae in 10 µm and 17 areolae in 10 µm.
Examined material: Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Catunda, Celso weir (UPCB: 78411, HUVA: 24535).
Species found in epilithic and epiphytic samples. Literature consulted: Buczkó et al. ( 2010Buczkó K, Ognjanova-Rumenova N & Magyari E (2010) Taxonomy, morphology and distribution of some Aulacoseira taxa in Glacial Lakes in the south Carpathian region. Polish Botanical Journal 55: 149-163.); Tremarin et al. ( 2013Tremarin PI, Ludwig TV & Torgan LC (2013) Morphological variation and distribution of the freshwater diatom Aulacoseira ambigua (Grunow) Simonsen in Brazilian continental environments. Iheringia Série Botânica 68: 139-157.).
Aulacoseira granulata var. granulata (Ehrenberg) Simonsen, Bacill. 2:58. 1979Simonsen R (1979) The diatom system: ideas on phylogeny. Bacillaria 2: 9-71.. Fig. 2c-f
Frustules cylindrical, united in straight filaments; terminal valves with one or two long spines; mantle ornamented with striae arranged parallel to the pervalvar axis; areolae rounded to rectangular; coarse, v-shaped sulcus not very pronounced. Diameter 5.7–7.7 μm; mantle height 11.6–21.3 μm; 8–12 striae in 10 μm and 8–10 areolae in 10 μm.
Examined material: Varjota, Araras weir (UPCB: 78392, HUVA: 24516). Sobral, Jaibaras weir (UPCB: 78395, HUVA: 24519). Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Sobral, Acaraú River (UPCB: 78407, HUVA: 24531). Meruoca, Sítio Cachoeira (UPCB: 78418, HUVA: 24542).
The species was found in epilithic, epiphytic and phytoplanktonic samples. Literature consulted: Krammer ( 1991Krammer K (1991) Morphology and taxonomy in some taxa of the genus Aulacoseira Thwaites (Bacillariophyceae). II. Taxa in the A. granulata-, italica- and lirata-groups. Nova Hedwigia 53: 477-496.); Houk ( 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Aulacoseira granulata var. angustissima (O. Müller) Simonsen, Bacill., 2:58. 1979Simonsen R (1979) The diatom system: ideas on phylogeny. Bacillaria 2: 9-71.. Fig. 2g-l
Frustules cylindrical, united in predominantly straight filaments by small connecting spines, terminal valves with a long spine; mantle ornamented by obliquely striae (dextrorse pattern) in the connecting valves and parallel in the separation valves; areolae rounded; sulcus not pronounced. Diameter 3–4 µm; mantle height 10.6–13.3 µm; 16–17 striae in 10 µm and 18–20 areolae in 10 µm.
Examined material: Varjota, Araras weir (UPCB: 78392, HUVA: 24516). Sobral, Jaibaras weir (UPCB: 78395, HUVA: 24519). Sobral, Acaraú River (UPCB: 78404, 78407, HUVA: 24528, 24531). Catunda, Carmina weir and Celso weir (UPCB: 78409, 78410, HUVA: 24533, 24534).
Aulacoseira granulata var. angustissima was found mainly in epiphytic samples. Literature consulted: Krammer ( 1991Krammer K (1991) Morphology and taxonomy in some taxa of the genus Aulacoseira Thwaites (Bacillariophyceae). II. Taxa in the A. granulata-, italica- and lirata-groups. Nova Hedwigia 53: 477-496.); Houk ( 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Aulacoseira italica (Ehrenberg) Simonsen, Bacill. 2:60. 1979Simonsen R (1979) The diatom system: ideas on phylogeny. Bacillaria 2: 9-71.. Fig. 2m-q
Frustules cylindrical, united in straight filaments by conspicuous conneting spines; valve face flat with inconspicuous areolae ( Fig. 2m); mantle ornamented with striae parallel to the pervalvar axis; areolae rounded, delicate. Diameter 8.5–10.6 µm; mantle height 12.6–17.4 µm; 22–24 striae in 10 µm and 18–22 areolae in 10 µm.

a-q. Aulacoseira species – a-b. Aulacoseira ambigua, in valve and girdle views, respectively; c-f. Aulacoseira granulata var. granulata; g-l. Aulacoseira granulata var. angustissima; m-q. Aulacoseira italica – m. valvar view; n-q. girdle view. Scale bar: 10 μm.
Examined material: Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Catunda, Celso weir (UPCB: 78411, HUVA: 24535).
Species found in epiphytic and epilithic samples. Literature consulted: Crawford et al. ( 2003Crawford RM, Likhoshway YV & Jahn R (2003) Morphology and identity of Aulacoseira Italica and typification of Aulacoseira (Bacillariophyta). Diatom Research 18: 1-19.); Houk ( 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.).
Bacillariophytina Medlin et Kaczmarska
Mediophyceae (Jousé et Proshkina-Lavrenko)
Medlin et Kaczmarska
Stephanodiscales Nikolaev et Harwood
Stephanodiscaceae Makarova
Cyclotella (Kützing) Brébisson
Cyclotella atomus Hustedt, Arch. Hydrobiol. 15:143, pl. 9, figs 1- 4. 1937Hustedt F (1937) Systematische und ökologische Untersuchungen über die Diatomeen-Flora von Java, Bali und Sumatra nach dem Material der Deutschen Limnologischen Sunda-Expedition. Archiv für Hydrobiologie (Supplement) 15: 131-177.. Fig. 3a-j
Valves circular, ornamented by marginal radiate striae separated by costae; marginal fultoportula ring, with fultoportulae distributed every 3 or 4 striae; one rimoportula located between two marginal fultoportulae; one subcentral fultoportula. Diameter 5–6.5 μm; pervalvar axis 3 μm; 8–14 striae in 10 μm and 3.3–3.8 marginal fultoportulae in 10 μm.
Examined material: Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Massapê, temporary pond near to Acaraú River (UPCB: 78399, HUVA: 24523). Sobral, Acaraú River (UPCB: 78403, 78407, 78408, HUVA: 24527, 24531, 24532).
In SEM: valve surface is slightly undulated in the central area ( Fig. 3h) and striae irregularly distributed from marginal valve face to mantle with delicate round poroids; marginal fultoportula ring located at the valve face/mantle junction, occurring at an interval of every three or four costae ( Fig.3i,j). Externally, fultoportulae and rimoportula openings are simple pores. Internally, striae are not alveolate ( Fig. 3j); the subcentral fultoportula have three satellite pores and marginal fultoportulae have two satellite pores in radial position; one sessile marginal rimoportula with a slightly oblique labiate opening ( Fig. 3j).

a-j. Cyclotella atomus – a-g. valves in LM; h-j. frustules and valves in SEM – h. frustule in girdle view showing marginal fultoportula external opening (arrow); i. external valve face view; j. internal view of the valve, marginal fultoportulae with two satellite pores (arrowhead), subcentral fultoportula with three satellite pores and marginal rimoportula with obliquely labiate opening (dark arrow). Scale bars: a-g = 10 μm; h = 2 μm; i-j = 1 μm.
Populations of Cyclotella atomus in northwest Ceará follow the morphometric variation described in the literature (diameter 3.5–8.5 μm), but the striae density showed lower values (12–20 in 10 μm; Tanaka 2007Tanaka H (2007) Taxonomic studies of the genera Cyclotella (Kützing) Brébisson, Discostella Houk et Klee and Puncticulata Håkansson in the Family Stephanodiscaceae Glezer et Makarova (Bacillariophyta) in Japan. Bibliotheca Diatomologica, Band 53. J Cramer, Berlim. 204p.; Houk et al. 2010 Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.). However, Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.) have also registered lower striae density (9–16 in 10 μm) in specimens from Northeastern Brazil. Cyclotella cryptica Reimann, Lewin et Guillard mainly differs from C. atomus due to the marginal fultoportulae located at an interval of every one or two costae ( Houk et al. 2010 Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.).
The species occurred in epiphytic, epilithic and phytoplanktonic samples. Literature consulted: Tanaka ( 2007Tanaka H (2007) Taxonomic studies of the genera Cyclotella (Kützing) Brébisson, Discostella Houk et Klee and Puncticulata Håkansson in the Family Stephanodiscaceae Glezer et Makarova (Bacillariophyta) in Japan. Bibliotheca Diatomologica, Band 53. J Cramer, Berlim. 204p.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.); Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.).
Cyclotella cryptica Reimann, Lewin et Guillard. Phycol. 3:82, figs. 4- 11. 1963Reimann BEF, Lewin JMC & Guillard RRL (1963) Cyclotella cryptica, a new brackish-water diatom species. Phycologia 3: 75-84.. Fig. 4a-k
Valves circular, ornamented by marginal radiate striae separated by costae; marginal fultoportulae ring, fultoportulae distributed in an interval of every one or two costae and always associated to the costa; one sessile rimoportula located between two marginal fultoportulae; one subcentral fultoportula. Diameter 6–12 μm; pervalvar axis 5.9 μm; 6–8 striae in 10 μm and 2–4.5 marginal fultoportulae in 10 μm. Examined material: Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Massapê, temporary pond near the Acaraú River (UPCB: 78399, HUVA: 24523). Sobral, Acaraú River (UPCB: 78403, 78406, 78407, 78408, HUVA: 24527, 24530, 24531, 24532). Taperuaba, lake in Pedra da Andorinha (UPCB: 78402, HUVA: 24526). Catunda, Carmina weirs (UPCB: 78409, HUVA: 24533) and Celso weirs (UPCB: 78410, HUVA: 24534). Varjota, Araras weir (UPCB: 78392, HUVA: 24516).
In SEM: internally, semi-open alveolate striae ( Fig. 4i-k); subcentral and marginal fultoportulae with three satellite pores around a long tube ( Fig. 4i-j); one marginal rimoportula with small labiate opening that is oriented obliquely lies on the costa ( Fig. 4j).
Cyclotella cryptica and C. meneghiniana Kützing are similar in valve diameter and number of marginal processes, but C. meneghiniana differs mainly due to the internally closed alveoli, which makes the separation between the central and marginal areas quite evident ( Houk et al. 2010 Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.). Moreover, it has already been observed that C. cryptica may resemble C. meneghiniana when found in low salinity (1.4) environments and show typical morphological characteristics of C. cryptica when salinity is > 4.3 ( Schultz 1971Schultz ME (1971) Salinity-related polymorphism in the brackish-water diatom Cyclotella cryptica. Canadian Journal of Botany 49: 1285-1289.).
The species occurred in epiphytic, epilithic, and phytoplanktonic samples. Literature consulted: Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.); Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.).
Cyclotella marina (Tanimura, Nagumo et M. Kato) Aké-Castillo, Okolodkov. et Ector, in Aké-Castillo et al. Nova Hedwigia Beih. 141: 267, figs 2- 9. 2012. Fig. 5a-h
Valves circular, delicate ornamentation, difficult to observe in MO; three to four marginal fultoportulae distant from each other; one rimoportula between two marginal fultoportulae; alveolate striae inconspicuous. Diameter 2.6–4.5 μm; 2–4 marginal fultoportulae in 10 μm.
Examined material: Sobral, Acaraú River (UPCB: 78403, 78405, 78408, HUVA: 24527, 24529, 24532).
In SEM: internal valve surface flat; striae radiate, delicate, extending toward the central region; alveoli absent and marginal fultoportulae surrounded by two satellite pores, arranged at the face/mantle junction and one pedunculated marginal rimoportula ( Fig. 5h).
Cyclotella marina differs from C. atomus due to less developed alveolate striae and the absence of central fultoportula ( Tanimura et al. 2004 Tanimura Y, Nagumo T & Kato M (2004) A new variety of Cyclotella atomus from Tokyo Bay, Japan, C. atomus var. marina var. nov. Bulletin of the National Science Museum Series C (Geology and Paleontology) 30: 5-11.). Also, the literature states the preference of C. marina for coastal, high-nutrient marine environments ( Aké-Castillo et al. 2012 Aké-Castillo JA, Okolodkov YB, Espinosa-Matías S, Merino-Virgilio FC, Herrera-Silveira JA & Ector L (2012) Cyclotella marina (Tanimura, Nagumo et Kato) Aké-Castillo, Okolodkov et Ector comb. et stat. nov. (Thalassiosiraceae): a bloom-forming diatom in the southeastern Gulf of Mexico. Nova Hedwigia, Beiheft (Suppl.) 141: 263-274.; Hevia- Orube et al. 2015 Hevia-Orube J, Orive E, David H, Díez A, Laza-Martínez A, Miguel I & Seoane S (2015) Molecular and morphological analyses of solitary forms of brackish Thalassiosiroid diatoms (Coscinodiscophyceae), with emphasis on their phenotypic plasticity. European Journal of Phycology 51: 1-20.). However, the species has been registered in freshwater environments, which implies that it has a wide distribution ( Cavalcante et al. 2013 Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.; Genkal & Yarmoshenko 2013Genkal SI & Yarmoshenko LP (2013) Centric Diatoms (Bacillariophyta) of the cooling pond of the Khmelnitskiy Nuclear Power Station (Ukraine). Hydrobiological Journal 49: 51-63.; Genkal & Okhapkin 2013Genkal SI & Okhapkin AG (2013) Centric Diatoms (Centrophyceae) of the lower reaches of the Oka river (Russian Federation). Hydrobiological Journal 49: 41-57.; Genkal & Bilous 2015Genkal SI & Bilous OP (2015) Centric Diatoms (Centrophyceae) of the lower portion of the Southern Bug river (Ukraine). International Journal on Algae 17: 339-350.). Cyclotella marina was registered in Brazil by Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.) in riverine waters of Bahia state, Northeastern Brazil. According to Tanimura et al. ( 2004Tanimura Y, Nagumo T & Kato M (2004) A new variety of Cyclotella atomus from Tokyo Bay, Japan, C. atomus var. marina var. nov. Bulletin of the National Science Museum Series C (Geology and Paleontology) 30: 5-11.) and Chung et al. ( 2010Chung MH, Yoon WD & Lee JB (2010) Morphological description of Cyclotella atomus var. marina (Bacillariophyceae): newly reported in Korean waters Algae 25: 57-64.), it is a metaphytic species occurring in both plankton and epiphyton. The present study is the second record of C. marina in freshwater systems in Brazil.
The species was registered exclusively in Acaraú River, influenced by domestic sewage, in epiphytic, epipsamic, and phytoplanktonic samples. Literature consulted: Aké-Castillo et al. ( 2012Aké-Castillo JA, Okolodkov YB, Espinosa-Matías S, Merino-Virgilio FC, Herrera-Silveira JA & Ector L (2012) Cyclotella marina (Tanimura, Nagumo et Kato) Aké-Castillo, Okolodkov et Ector comb. et stat. nov. (Thalassiosiraceae): a bloom-forming diatom in the southeastern Gulf of Mexico. Nova Hedwigia, Beiheft (Suppl.) 141: 263-274.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Cyclotella meduanae Germain, Flore des Diatomées. p. 36, pl. 8, fig. 28, pl. 154, figs. 4, 4a. 1981Germain H (1981) Flore des diatomées Diatomophycées eaux douces et saumâtres du Massif Armoricain et des contrées voisines d’Europe occidentale. Collection "Faunes et Flores Actuelles". Société Nouvelle des Editions Boubée, Paris. 444p.. Fig. 5i
In SEM: internal view, circular valve ornamented by marginal radiate striae with delicate areolae, separated by costae; striae are not alveolate; subcentral fultoportulae absent; marginal fultoportulae ring surrounded by three satellite pores, located at the face/valve mantle junction, at every two or three striae; one marginal rimoportula located between two fultoportulae on the costae, obliquely oriented. Diameter 5.9 μm; 9.5 striae in 10 μm and 4.3 marginal fultoportulae in 10 μm.
Examined material: Sobral, Acaraú River (UPCB: 78408, HUVA: 24532). Catunda, Carmina weir (UPCB: 78409, HUVA: 24533).
The only specimen found during SEM preparations is consistent with descriptions by Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.) and Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.). Cyclotella katiana Sala et Ramírez, which was proposed in Colombia ( Sala & Ramírez 2008Sala S & Ramírez JJ (2008) Cyclotella katiana sp. nov. from La Reina Swamp, Parque Nacional Natural los Katíos, Colombia. Diatom Research 23: 147-167.), was recently considered synonymous with C. meduanae based on their overlapping diacritical characteristics ( Genkal 2014Genkal SI (2014) Morphology, taxonomy, ecology and distribution of Cyclotella meduanae Germain (Bacillariophyta). Nova Hedwigia Beiheft 143: 127-140.). Cyclotella meduanae resembles C. cryptica but differs mainly in relation to the absence of the subcentral fultoportula in C. meduanae ( Houk et al. 2010 Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.).

a-k. Cyclotella cryptica – a-h. valves in LM; i-k. valves in SEM – i. internal valve view, detail of the subcentral fultoportula with three satellite pores (arrowhead); j. marginal fultoportula with three satellite pores (arrowhead), and detail of the sessile marginal rimoportula (dark arrow), and detail of the semi-open alveoli and ring of marginal fultoportulae; k. valve overview showing details of subcentral fultoportula and marginal rimoportula and fultoportula ring. Scale bars: a-h = 10 μm; k = 2 μm; i-j = 1 μm.

a-i. Cyclotella species – a-g. Cyclotella marina – valve view in LM; h. internal view of the valve in SEM, detail of the sessile marginal rimoportula (arrow). i. Cyclotella meduanae – internal valve view in SEM, detail of the marginal rimoportula between a costa and a marginal fultoportula (arrow). Scale bars: a-g = 10 μm; h-i = 1 μm.
The species occurred only in plankton in epiphyton. Literature consulted: Tanaka ( 2007Tanaka H (2007) Taxonomic studies of the genera Cyclotella (Kützing) Brébisson, Discostella Houk et Klee and Puncticulata Håkansson in the Family Stephanodiscaceae Glezer et Makarova (Bacillariophyta) in Japan. Bibliotheca Diatomologica, Band 53. J Cramer, Berlim. 204p.); Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Cyclotella meneghiniana Kützing, Die Kies. Bacill. oder Diat., p. 50, pl. 30, fig. 68. 1844Kützing FT (1844) Die Kieselschaligen Bacillarien oder Diatomeen. Zu finden bei W. Köhne, Nordhausen. Pp. [i-vii], [1]-152, pls 1-30.. Fig. 6a-k
Valves circular, with evident separation between central and marginal areas, central area with tangential undulation; marginal striae alveolate; marginal fultoportula ring, with one fultoportula in each stria, sometimes absent; one rimoportula inserted between two marginal fultoportulae; one to two subcentral fultoportula. Diameter 7.9–18.4 μm; 7–8 striae in 10 μm and 4.9 marginal fultoportulae in 10 μm.
Examined material: Varjota, Araras weir (UPCB: 78392, HUVA: 24516). Granja, Gangorra weir (UPCB: 78393, 78394, HUVA: 24517, 24518). Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Massapê, temporary pond near the Acaraú River (UPCB: 78399, HUVA: 24523). Taperuaba, lake in Pedra da Andorinha and Olho d’água do Pajé (UPCB: 78402, 78400, HUVA: 24526, 24524). Sobral, Acaraú River (UPCB: 78403, 78404, 78406, 78407, 78408, HUVA: 24527, 24528, 24530, 24531, 24532). Catunda, Carmina and Celso weirs (UPCB: 78409, 78410, HUVA: 24533, 24534). Viçosa do Ceará, Quatiguaba River (UPCB: 78413, HUVA 24537).
In SEM: the external valve surface is slightly undulated in the central region, ornamented by granules; subcentral fultoportulae; marginal single or double spines positioned in line with each costa (Fig. 6i). Internal view with closed alveoli, marginal and subcentral short tube fultoportulae surrounded by three satellite pores ( Fig. 6j-k); one pedunculated marginal rimoportula between two marginal fultoportulae, with obliquely oriented labiate opening ( Fig. 6k).

a-k. Cyclotella meneghiniana (valve view in LM and SEM) – a-h. external view of valve surface in LM; i. external view of valve surface in SEM, detail of the valve surface, of the marginal spines; j. internal view, alveolate striae, closed alveolous (white arrow), subcentral fultoportula with three satellite pores (dark arrow); k. pedunculated marginal rimoportula (arrowhead) obliquely oriented, marginal fultoportula (dark arrow). Scale bars: a-h = 10 μm; i, j = 5 μm; k = 2 μm.
The species occurred in epiphytic, epilithic, and phytoplanktonic samples. Literature consulted: Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Discostella Houk et Klee
Discostella stelligera (Cleve et Grunow) Houk et Klee, Diat. Res. 19(2): 208. 2004. Fig. 7a-i
Valves circular, with convex central area, ornamented by short striae irregularly arranged in form of a rosette; marginal area occupies less than half of the valve surface; marginal alveolate striae, radial and regular in length; marginal ring of fultoportulae inconspicuous in LM. Diameter 5.9–12 µm and 7.5–11.5 striae in 10 µm.
Examined material: Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Granja, Gangorra weir (UPCB: 78393, HUVA: 24517). Sobral, Acaraú River (UPCB: 78403, 78409, HUVA: 24527, 24533).
In SEM: internal view with marginal fultoportulae located in line with striae and between costae, in the valve face/mantle junction, surrounded by two satellite pores; one small marginal rimoportula present between marginal fultoportulae ( Fig. 7i). The internal opening of the alveolate striae is shortened when it coincides with fultoportula or rimoportula. A hyaline area separates the marginal from the central striations.
Species found mainly in epiphytic samples. Literature consulted: Houk ( 2004Houk V & Klee R (2004) The stelligeroid taxa of the genus Cyclotella (Kutz.) Brébisson (Bacillariophyceae) and their transfer to the new genus Discostella gen. nov. Diatom Research 19: 203-228.); Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.); Tuji & Williams ( 2006Tuji A & Williams DM (2006) Type examination of Cyclotella wortereckii Hust. (Bacillariophyceae) with special attention to the position of its rimoportula. Bulletin of the National Science Museum Series B (Botany) 32: 15-17.); Guerrero & Echenique ( 2006Guerrero JM & Echenique RO (2006) Discostella taxa (Bacillariophyta) from the Río Limay basin (northwestern Patagonia, Argentina). European Journal of Phycology 41: 83-96.).
Discostella woltereckii (Hustedt) Houk et Klee, Diat. Res. 19(2): 223. 2004. Fig. 7j-o
Valves circular, flat to a moderately convex central area, with two ornamentation patterns: elongated marginal striae, of irregular length, which extends to the valve center ( Fig. 7j-m), or irregular marginal striae occupying more than one half of the valve, separated from the central region, which presents short radiated rosette-shaped striation, ( Fig. 7n-o); marginal ring of fultoportulae inconspicuous in LM. Diameter 4.8–5 μm; 8.5 striae in 10 μm.

a-o. Discostella species – a-i. Discostella stelligera – a-h. valves in LM; i valve in internal view, SEM; j-o. Discostella wortereckii – valves in LM. Scale bars: a-h, j-o = 10 μm; i = 2 μm.
Examined material: Varjota, Araras weir (UPCB: 78392, HUVA: 24516).
Discostella woltereckii var. minor Öberg, Risberg et Stabell differs from the typical species due to the smaller valve diameter (1.9–4 μm) and the dichotomously arranged valve face ( Öberg et al. 2009 Öberg H, Risberg J & Stabell B (2009) Morphology, valve ultrastructure and stratigraphical variability of Discostella taxa in a tropical crater lake, northern Tanzania. Diatom Research 24: 341-356.). Discostella guslyakovyi Genkal, Bondarenko et Popovskaya also differs in diameter (2.8–5.7 μm), valve contour, and non-tubular marginal fultoportulae ( Genkal et al. 2007 Genkal SI, Bondarenko NA & Popovskaya GI (2007) New representative of the genus Discostella Houck et Klee (Bacillariophyta) from the Eastern Baikal area (Russia). International Journal on Algae 9: 359-364.).
Similarities are notable between D. pseudostelligera (Hustedt) Houk et Klee and D. woltereckii. Both have irregular marginal striations, long tubular marginal fultoportulae, and may present a central area ornamented by short striae arranged in a rosette shape ( Houk et al. 2010 Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.). However, according to Guerrero & Echenique ( 2006Guerrero JM & Echenique RO (2006) Discostella taxa (Bacillariophyta) from the Río Limay basin (northwestern Patagonia, Argentina). European Journal of Phycology 41: 83-96.), D. pseudostelligera exhibits a broad hyaline ring occupying about half of the valve diameter, located between the central and the marginal area, while in D. woltereckii the central area is very small, sometimes reduced to an isolated stria.
The population in Ceará is similar to that presented by Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.) fig.19, pl.354, Hustedt ( 1942Hustedt F (1942) Süßwasser-Diatomeen des indomalayischen Archipels und der Hawaii-Inseln. Nach dem Material der Wallacea-Expedition. Internationale Revue der gesamten Hydrobiologie und Hydrographie 42: 1-252.) fig.25, pl.324 for D. woltereckii type material, and also Huber-Pestalozzi ( 1942, fig. 488A, pl. CXVIIIHuber-Pestalozzi G (1942) Diatomeen. In: Thienimann A (ed.) Das phytoplankton des susswassers, systematik und biologie. Stuttgart E. Schweizerbartsche Verlagsbuchhandlung 2: 367-549.). When analyzing the type material of both taxa, Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.) stated that, although the morphology can sometimes overlap, making identification inaccurate, the separation of the taxa can be related to ecological differences; D. pseudostelligera is mainly a species of temperate regions, while D. woltereckii occurs preferentially in tropical zones. Morphological and ecological features led us to assume that the population in Ceará corresponds better with the characterization of D. woltereckii.
The studied population was recorded in epiphytic samples. Literature consulted: Houk et al. ( 2010Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.); Guerrero & Echenique ( 2006Guerrero JM & Echenique RO (2006) Discostella taxa (Bacillariophyta) from the Río Limay basin (northwestern Patagonia, Argentina). European Journal of Phycology 41: 83-96.).
Thalassiosirales Glezer et Makarova
Thalassiosiraceae M. Lebour
Conticribra Stachura-Suchoples et D.M. Williams Conticribra weissflogii (Grunow) Stachura- Suchoples et Williams, Eur. J. Phycol. 44(4): 482. 2009. Fig. 8a-m
Valves circular with a flat surface; striae delicate, irregularly oriented; marginal fultoportulae ring located in the face/mantle junction, with conspicuous external tubes; one pronounced rimoportula interrupting the ring of marginal fultoportulae. Diameter 19.8–32 μm; marginal fultoportulae 5.5–7.6 in 10 μm; central fultoportulae 5–6 in 10 μm.
Examined material: Graça, Belizário waterfall (UPCB: 78398, HUVA: 24522). Sobral, Acaraú River (UPCB: 78403, 78406, 78408, HUVA: 24527, 24530, 24532).
In SEM: external valve surface ornamented with granules ( Fig. 8h-j) and areolate striae with an irregular continuous radial pattern. Marginal ring of fultoportulae with long external tubes, central fultoportulae with moderately elongated tubes ( Fig. 8i-j). Rimoportula with an elongated external tube, slightly longer than those of fultoportulae ( Fig 8i). For internal view, marginal and central fultoportulae are present as small tubes with four satellite pores ( Fig. 8k-m). Rimoportula with a large labial opening, short pedunculated, arranged radially ( Fig. 8l).

a-m. Conticribra weissflogii (valve view in LM and SEM) – a-g. valve view in LM, flat surface and delicate stretch marks; h-j. external view of valve surface in SEM – h. external valve surface ornamented with granules, marginal ring of fultoportulae with long external tubes; i. details of marginal fultoportulae and rimoportulae openings (arrow); j. central fultoportulae openings; k-m. internal valve view in SEM – k. internal surface of the valve without pronounced ornamentation, internal view of the ring of marginal fultoportulae interrupted by a rimoportula; l. details of large sessile rimoportula; m. central fultoportulae with for satellite pores surrounding a short tube. Scale bars: a-g = 10 μm; h-j = 5 μm; k-m = 2 μm.
The species occurred in epilithic, epiphytic, and phytoplanktonic samples. Literature consulted: Stachura-Suchoples & Williams DM ( 2009Stachura-Suchoples K & Williams DM (2009) Description of Conticribra tricircularis, a new genus and species of Thalassiosirales, with a discussion on its relationship to other continuous cribra species of Thalassiosira Cleve (Bacillariophyta) and its freshwater origin. European Journal of Phycology 44: 477-486.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Eupodiscales Bessey Eupodiscaceae Ralfs
Pleurosira (Meneghini) Trevisan
Pleurosira laevis (Ehrenberg) Compère var. laevis, Bacill., 5: 177-178, fig. 1-17, 20, 39. 1982. Fig. 9a-d; 10a-f

Subcircular to elliptical valves; valve face flat, ornamented by radial striae with delicate round areolae; two large ocelli located at the valve margin, opposite to each other, ornamented by delicate poroids; two rimoportulae located near the center of the valve. Larger diameter 38.1–45.5 μm; smaller diameter 30.9–47.2 μm; 10–14 areolae in 10 μm.
Examined material: Ipu, Bica do Ipu (UPCB: 78396, HUVA: 24520). Sobral, Acaraú River (UPCB: 78403, HUVA: 24527). Viçosa do Ceará, Quatiguaba River (UPCB: 78412, 78413, HUVA: 24536, 24537).
In SEM: valve surface is ornamented by irregular starry spines mostly concentrated in the central region and close to the ocelli ( Fig. 10i-k); rimoportulae with an external opening as a simple slit ( Fig. 10h) and internally as a sessile labiate opening ( Fig. 10g).

a-f. Pleurosira laevis var. laevis (valve view in SEM) – a. internal view of the valve, two rimoportulae located near the center of the valve; b. detail of the rimoportula opening in internal view (arrow); c. external view of the rimoportula opening; d. external valve surface ornamented with small spines irregularly spaced; e. external view of ocellus; f. detail of the spines, SEM. Scale bars: a-b,d = 10 μm; f = 5 μm; c,e = 2 μm.
Pleurosira laevis var. paludosa (Tempère et Peragallo ex Forti) Compère, differs from the typical variety as it shows rimoportulae closer to the central area ( Compère 1982Compère P (1982) Taxonomic revision of the diatom genus Pleurosira (Eupodiscaceae). Bacillaria 5: 165-190.). Pleurosira socotrensis (Kitton) Compère differs due to its elliptical valves and irregularly arranged striae in the central region of the valve ( Compère 1982Compère P (1982) Taxonomic revision of the diatom genus Pleurosira (Eupodiscaceae). Bacillaria 5: 165-190.; Ludwig et al. 2004 Ludwig TAV, Flores TL, Moreira-Filho H & Veiga LAS (2004) Inventário florístico das diatomáceas (Ochrophyta) de lagoas do Sistema Hidrológico do Taim, Rio Grande do Sul, Brasil: Coscinodiscophyceae. Iheringia, Série Botânica 59: 97-106.; Karthick & Kociolek 2011Karthick B & Kociolek JP (2011) Four new centric diatoms (Bacillariophyceae) from the Western Ghats, South India. Phytotaxa 22: 25-40.).
Population recorded mainly in epilithic and epiphytic samples. Literature consulted: Compère ( 1982Compère P (1982) Taxonomic revision of the diatom genus Pleurosira (Eupodiscaceae). Bacillaria 5: 165-190.); Joh ( 2010Joh G (2010) Algal flora of Korea. Chrysophyta, Bacillariophyceae: centrales. Freshwater diatoms I. National Institute of Biological Resources 3: 1-161.); Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.).
Orthoseirales Crawford
Orthoseiraceae Crawford
Orthoseira Thwaites
Orthoseira roeseana (Rabenhorst) Pfitzer, Bot. Abh. 1(2): 134. 1871Pfitzer E (1871) Untersuchungen über bau und entwicklung der bacillariaceen (Diatomaceen). In: von Hanstein JLER (ed.) Botanische abhandlungen aus dem Gebiet der Morphologie und Physiologie. Vol. 1. A. Marcus, Bonn. 189p.. Fig. 11a-h
Cylindrical frustule in girdle view, united by inconspicuous spines; mantle ornamented by striae parallel to the pervalvar axis, little pronounced constriction (stricter region in the mantle) ( Fig. 11a-c); valve surface slightly wavy, with scattered punctuations, striae radial, conspicuously areolate; central area with three carinoportulae ( Fig. 11e). Diameter 8.7–26.4 μm; mantle height 25.7–35.6; 15–17 striae in 10 μm; 18–19 areolae in 10 μm.
Examined material: Ipu, Bica do Ipu (UPCB: 78396, HUVA: 24520). Ibiapina, Bica do Pajé (UPCB: 78397, HUVA: 24521). Ubajara, Sítio São Luis (UPCB: 78414, HUVA: 24538).
In SEM: carinoportulae occluded internally ( Fig. 11g) and externally, presence of spines ( Fig. 11h).

a-h. Orthoseira roeseana – a-f. valves in LM – a-c. girdle views, observe the valve constrictions (arrow); d-f. valve views; g-h. valves in SEM – g. detail of carinoportulae in internal view; h. external view. Scale bars: a-f = 10 μm; g-h = 5 μm.
In Brazilian studies, specimens of Orthoseira roesena are presented by Landucci & Ludwig ( 2005Landucci M & Ludwig TAV (2005) Diatomáceas de rios da Bacia Hidrográfica Litorânea, PR, Brasil: Coscinodiscophyceae e Fragilariophyceae. Acta Botanica Brasilica 19: 345-357., fig. 1), Brassac et al. ( 1999Brassac NM, Atab DR, Landucci M, Visinoni ND & Ludwig TAV (1999) Diatomáceas cêntricas de rios da região de abrangência da usina hidrelétrica de Salto Caxias, PR (Bacia do Rio Iguaçu) Acta Botanica Brasilica 13: 277-289., fig. 29), and Ferrari & Ludwig ( 2007Ferrari F & Ludwig TAV (2007) Coscinodiscophyceae, Fragilariophyceae e Bacillariophyceae (Achnanthales) dos rios Ivaí, São João e dos Patos, bacia hidrográfica do Rio Ivaí, município de Prudentópolis, PR, Brasil. Acta Botanica Brasilica 21: 421-441., figs 7 and 8), while Orthoseira dendroteres (Ehrenberg) Genkal et Kulikovskiy is presented by Nardelli et al. ( 2014Genkal SI (2014) Morphology, taxonomy, ecology and distribution of Cyclotella meduanae Germain (Bacillariophyta). Nova Hedwigia Beiheft 143: 127-140., fig. 13). When comparing the illustrations and descriptions of these studies, they likely correspond to the same taxon. The delimitation between O. dendroteres and O.roeseana becomes difficult due to the vast morphological variation of both taxa ( Houk 1993Houk V (1993) Some morphotypes in the Orthoseira roeseana complex. Diatom Research 8: 385-402., 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.; Spaulding & Kociolek 1998Spaulding SA & Kociolek JP (1998) The diatom genus Orthoseira: ultrastructure and morphological variation in two species from Madagascar with comments on nomenclature in the Genus. Diatom Research 13: 133-147.). The populations in this study exhibited considerable morphological variation in terms of valve diameter, overlapping with the characteristics of both species. Therefore, we opted to adhere to the broad taxonomic concept of O. roeseana found in Houk ( 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.). Gargas et al. ( 2018Gargas CB, Theriot EC, Ashworthb MP & Johansen JR (2018) Phylogenetic analysis reveals that the ‘radial centric’ diatom Orthoseira Thwaites (Orthoseiraceae, Bacillariophyta) is a member of a ‘multipolar’ diatom lineage. Protist 169: 803-825.) comment on the need for more studies to resolve the taxonomy and the typification of O. dendroteres and O. roeseana.
Populations found in periphyton in epilithic of humid subaerial samples. Literature consulted: Houk ( 1993Houk V (1993) Some morphotypes in the Orthoseira roeseana complex. Diatom Research 8: 385-402., 2003Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.).
Terpsinoaceae Ralfs in Pritchard
Terpsinoë Ehrenberg
Terpsinoë musica Ehrenberg, Abh. Akad. Wiss. Berl., p. 425, pl. 3, fig. IV.1, pl. 3, fig. VII. 30. 1841 ( 1843Ehrenberg CG (1843) Verbreitung und Einfluss des mikroskopischen Lebens in Süd- und Nord-Amerika. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1841: 291-445.). Figs. 12a-q; 13a-h
Frustules rectangular in girdle view with transapical bars resembling musical notes and short pseudoseptum near the edge of valve mantle; bipolar, elongated valves, generally with three marginal undulations of nearly equal size, but smaller valves showed one ( Fig. 12i-m) to three marginal undulations ( Fig. 12a-g); valve ends rostrate to subcapitate with pseudocelli; transapical costae between each undulation and near to the valve ends (less developed); coarse areolae irregularly arranged on the surface of the valve; one or three ( Fig. 12a) subcentral rimoportula located in the central undulation. Length 61.7–137.9 µm; width 32.8–42.2 µm; pervalvar axis 74.5–107.9 µm and 8–10 areolae in 10 µm.

a-q. Terpsinoë musica (valves in LM) – a-m. valve view; n-q. girdle view. Scale bars: 10 μm.
Examined material: Ipu, Bica do Ipu (UPCB: 78396, HUVA: 24520).
In SEM: internal view shows a small rimoportula in form of a crack ( Fig. 13c) or a closed lip ( Fig. 13d), and the rimoportula external view shows a slit shape ( Fig. 13e).

a-h. Terpsinoë musica (valves in SEM) – a-b. internal valve view of the valve, observe the septa; c. internal view of the rimoportula, valve with less undulations; d. internal view of the rimoportula, valve with three undulations; e. external view of the rimoportula; f-g. valve apices of one or bi undulated valves; h. valve apice of triundulated valve. Scale bars: a = 20 μm; b,h = 10 μm; d-g = 5 μm; c. 2 μm.
The wide morphological variation of T. musica populations was previously recorded in the literature ( Schmidt 1812–1899Schmidt A (1812-1899) Verzeichniss der. In: A. Schmidt's Atlas der Diatomaceenkunde: Heft 1-36 (Serie I-III) abgebildeten Arten und benannten Varietäten nebst den mit angeführten Synonymen. Topics Diatoms. O.R. Reisland, Leipzig. Pp. 350.; Luttenton et al. 1986 Luttenton MR, Pfiester LA & Timpano P (1986) Morphology and growth habitat of Terpsinoe musica Ehr. (Bacillariophyceae). Castanea, Journal of the Southern Appalachian Botanical Club. Morgantown 51: 175-182.; Jiménez et al. 2017 Jiménez JA, Magos YB & Camarillo VHS (2017) Morphological and environmental characterization of Terpsinoë musica (Biddulphiaceae, Bacillariophyceae) in tropical streams from Mexico. Diatom Research 32: 185-193.; et al. 2005 Metzeltin D, Lange-Bertalot H & García-Rodríguez F (2005) Diatoms of Uruguay, compared with other taxa from South America and elsewhere. Iconographia Diatomologica (H. Lange-Bertalot, ed.). Vol. 15. Koeltz Scientifc Books, Stuttgart. 736p.; Metzeltin & Lange-Bertalot 2007Metzeltin D & Lange-Bertalot H (2007) Tropical Diatoms of South America II. Special remarks on biogeographic disjunction. Iconographia Diatomologica (H. Lange-Bertalot, ed.). Vol. 18. Koeltz Scientifc Books, Stuttgart. 877p.). However, documentation of specimens with only one central undulation is rare. Wu ( 2013Wu S (2013) Terpsinoë musica. Diatoms of North America. Available at < https://diatoms.org/species/terpsinoe_musica>. Access on 23 June 2020.
https://diatoms.org/species/terpsinoe_mu...
) described valves with one undulation in T. musica specimens, stating that the number of undulations decreases in smaller valves. Metzeltin & Lange-Bertalot ( 2007Metzeltin D & Lange-Bertalot H (2007) Tropical Diatoms of South America II. Special remarks on biogeographic disjunction. Iconographia Diatomologica (H. Lange-Bertalot, ed.). Vol. 18. Koeltz Scientifc Books, Stuttgart. 877p.) illustrated “ Terpsinöe (? nov.) spec.” with frustules containing a single undulation (pl. 296:5– 12), demonstrating uncertainties in identification. In our material, all valves were found in the same population (see Examined material), including intermediate forms between one and two marginal undulations. Our observations suggest that these forms correspond to the same taxon. Variability in morphology and symmetry may be the result of cell reduction or initial cell formation ( Cox 2014Cox EJ (2014) Diatom identification in the face of changing species concepts and evidence of phenotypic plasticity. Journal of Micropalaeontology 33: 111-120.), as in the interrupted projections of the bipolar diatom Hydrosera ( Cox 2013Cox EJ (2013) Recognising and defining taxonomic boundaries in ‘well-known’ freshwater diatoms and its relevance to water quality evaluation. Diatomededelingen 37: 17-26.). Functional aspects, such as nutrient availability and adaptation to the environment, also provoke significant morphological variability in populations ( Jiménez et al. 2017 Jiménez JA, Magos YB & Camarillo VHS (2017) Morphological and environmental characterization of Terpsinoë musica (Biddulphiaceae, Bacillariophyceae) in tropical streams from Mexico. Diatom Research 32: 185-193.).
Small specimens with a single central undulation from the study population are similar to Terpsinoë petitiana (Leuduger-Fortmorel) Hendey found in marine samples from West Africa ( Leuduger-Fortmorel 1898Leuduger-Fortmorel G (1898) Diatomées marines da costa ocidental de África. Saint-Brieuc Francisque Guyon, Saint-Brieuc. 41p.) and the Galapagos ( Hendey 1972Hendey NI (1972) Some marine diatoms from the Galapagos Islands. Nova Hedwigia 22: 371-422.). The possibility remains that T. musica and T. petitiana are conspecifics, not only because of their phenotypic plasticity, but also because T. musica presents a wide ecological distribution, occurring in fresh and brackish water, as well as marine environments ( Round et al. 1990 Round FE, Crawford RM & Mann DG (1990) The diatoms: biology and morphology of the genera. Cambridge University Press, Cambridge. 747p.). In this study, specimens of T. musica were found on bryophytes growing on rocks in a humid subaerial environment, for which we documented valves with one to three central undulations (Fig. 11a-m).
Tuji ( 2018Tuji A (2018) A new freshwater diatom, Terpsinoë muninensis sp. nov., from the Ogasawara Islands, Japan. Memoirs of the National Science Museum (Tokyo) 52: 5-15.) describes T. muninensis, a species very similar to T. musica, as endemic to the freshwater of the North Pacific islands. The author differentiates T. muninensis as it presents less silicified apices and interrupted pseudosepta, which do not form musical notes, as is characteristic of T. musica ( Ehrenberg 1843Ehrenberg CG (1843) Verbreitung und Einfluss des mikroskopischen Lebens in Süd- und Nord-Amerika. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1841: 291-445.). However, this characteristic seems to vary within the same population. In the samples from Ceará, the pseudosepta present as well silicified ( Fig. 12d-e), with a low level of silicification ( Fig. 12a-c,f), or are not visible ( Fig. 12n-q). In the population described by Jiménez et al. ( 2017Jiménez JA, Magos YB & Camarillo VHS (2017) Morphological and environmental characterization of Terpsinoë musica (Biddulphiaceae, Bacillariophyceae) in tropical streams from Mexico. Diatom Research 32: 185-193.), Figs. 8 and 11 show well-developed pseudosepta, which is not seen in Fig. 10. The authors also state that T. musica would have a median marginal undulation greater than the others, differing from those of equal size in T. muninensis. However, Jiménez et al. ( 2017Jiménez JA, Magos YB & Camarillo VHS (2017) Morphological and environmental characterization of Terpsinoë musica (Biddulphiaceae, Bacillariophyceae) in tropical streams from Mexico. Diatom Research 32: 185-193.) illustrate ( Fig. 8) T. musica with marginal undulation of the same size and apices with well-developed pseudosepta. Tuji ( 2018Tuji A (2018) A new freshwater diatom, Terpsinoë muninensis sp. nov., from the Ogasawara Islands, Japan. Memoirs of the National Science Museum (Tokyo) 52: 5-15.) states that differences in the molecular sequence support the existence of two taxa, but also recommends further studies on the morphological and molecular phylogenetic variability of the two species. Also, some more representatives of the genus Terpsinoë have sequences deposited at GenBank that should be used by the author for a better comparison and a closer representation of the genus phylogeny.
In light of the above discussion, we believe that the Ceará population fits the concept of T. musica, a taxon with significant morphological plasticity.
Population found in periphyton of humid subaerial samples, associated with bryophytes. Literature consulted: Metzeltin et al. ( 2005Metzeltin D, Lange-Bertalot H & García-Rodríguez F (2005) Diatoms of Uruguay, compared with other taxa from South America and elsewhere. Iconographia Diatomologica (H. Lange-Bertalot, ed.). Vol. 15. Koeltz Scientifc Books, Stuttgart. 736p.); Wu ( 2013Wu S (2013) Terpsinoë musica. Diatoms of North America. Available at < https://diatoms.org/species/terpsinoe_musica>. Access on 23 June 2020.
https://diatoms.org/species/terpsinoe_mu...
); Jiménez et al. ( 2017Jiménez JA, Magos YB & Camarillo VHS (2017) Morphological and environmental characterization of Terpsinoë musica (Biddulphiaceae, Bacillariophyceae) in tropical streams from Mexico. Diatom Research 32: 185-193.).
Most taxa were rare (6) or sporadic (7). Only two species were frequent: Cyclotella meneghiniana, occurring in 16 of the 24 sampling points, and Cyclotella cryptica which occurred in 10 sampling points. Furthermore, only C. meneghiniana occurred in all hydrographic basins, showing its wide geographic distribution in the region. In general, C. meneghiniana is frequent and abundant in rivers and lakes, under different trophic conditions ( Kiss et al. 2012 Kiss KT, Klee R, Ector L & Ács É (2012) Centric diatoms of large rivers and tributaries in Hungary: morphology and biogeographic distribution. Acta Botanica Croatica 71: 311-363.), and is particularly common in shallow and nutrient-rich waters ( Houk et al. 2010 Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.). The species has been classified as an eutrophic indicator for highly fertilized waters with high conductivity ( Joh 2010Joh G (2010) Algal flora of Korea. Chrysophyta, Bacillariophyceae: centrales. Freshwater diatoms I. National Institute of Biological Resources 3: 1-161.).
The taxa that occurred in at least three of the studied hydrographic basins were: Cyclotella atomus, C. cryptica, and Discostella stelligera. Furthermore, it was observed that there was no significant difference in richness among different substrates, but rather in different environments, being higher in rivers and lower in puddles and dams.
Fifteen taxa were identified, with 12 new occurrences for the northwest region of Ceará. Aulacoseira ambigua (cited as Melosira ambigua (Grunow) O.Müller), Aulacoseira granulate var. granulata [cited as Melosira granulata (Ehrenberg) Ralfs], and Cyclotella meneghiniana were previously registered by Patrick ( 1940aPatrick R (1940a) Diatoms of Northeastern Brazil. Part I. Coscinodiscaceae, Fragilariaceae and Eunotiaceae. Proceedings of the Academy of Natural Sciences of Philadelphia 92: 191-226.) in the Jaibaras weir, in the municipality of Sobral. Discostella stelligera is registered for the first time in Northeastern Brazil, having been previously recorded in the South ( Ferrari & Ludwig 2007Ferrari F & Ludwig TAV (2007) Coscinodiscophyceae, Fragilariophyceae e Bacillariophyceae (Achnanthales) dos rios Ivaí, São João e dos Patos, bacia hidrográfica do Rio Ivaí, município de Prudentópolis, PR, Brasil. Acta Botanica Brasilica 21: 421-441.; Nardelli et al. 2014 Nardelli MS, Bueno NC, Ludwig TAV, Tremarin PI & Bartozek ECR (2014) Coscinodiscophyceae and Fragilariophyceae (Diatomeae) in the Iguaçu river, Paraná, Brazil. Acta Botanica Brasilica 28: 127-140.; Silva-Lehmkuhl et al. 2019 Silva-Lehmkuhl AM, Tremarin PI, Vercellino IS & Ludwig TAV (2019) Periphytic diatoms from an oligotrophic lentic system, Piraquara I reservoir, Paraná state, Brazil. Biota Neotropica 19: e20180568.), Southeast ( Morandi et al. 2006 Morandi LL, Ritter LMO, Moro RS & Bicudo CEM (2006) Criptógamos do Parque Estadual do Ipiranga, São Paulo, SP. Algas, 20: Coscinodiscophyceae. Hoehnea 33: 115-122.), and Central-west ( Da Silva et al. 2011 Da Silva WJ, Nogueira IS & Souza MGM (2011) Catálogo de diatomáceas da região Centro-Oeste brasileira. Iheringia Série Botânica 66: 61-86.) of the country. Discostella woltereckii is cited for the first time in Brazil; however, there are previous identifications of D. pseudostelligera for which conspecificity must be evaluated ( Brassac et al. 1999 Brassac NM, Atab DR, Landucci M, Visinoni ND & Ludwig TAV (1999) Diatomáceas cêntricas de rios da região de abrangência da usina hidrelétrica de Salto Caxias, PR (Bacia do Rio Iguaçu) Acta Botanica Brasilica 13: 277-289.; Morandi et al. 2006 Morandi LL, Ritter LMO, Moro RS & Bicudo CEM (2006) Criptógamos do Parque Estadual do Ipiranga, São Paulo, SP. Algas, 20: Coscinodiscophyceae. Hoehnea 33: 115-122.; Cavalcante et al. 2013 Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.; Faustino et al. 2016 Faustino SB, Fontana L, Bartozek ECR, Bicudo CEM & Bicudo DC (2016) Composition and distribution of diatom assemblages from core and surface sediments of a water supply reservoir in Southeastern Brazil. Biota Neotropica 16: e20150129.). The species complexes Discostella pseudostelligera/ D. woltereckii, Orthoseira roeseana/O. dendroteres and Terpsinoë musica/ T. muninensis require further attention and additional studies, including analyses of their ecology and lifecycle and assessments using molecular tools, to provide a better delimitation of each.
Cyclotella marina is registered for the second time in a Brazilian freshwater habitat. According to Aké-Castillo et al. ( 2012Aké-Castillo JA, Okolodkov YB, Espinosa-Matías S, Merino-Virgilio FC, Herrera-Silveira JA & Ector L (2012) Cyclotella marina (Tanimura, Nagumo et Kato) Aké-Castillo, Okolodkov et Ector comb. et stat. nov. (Thalassiosiraceae): a bloom-forming diatom in the southeastern Gulf of Mexico. Nova Hedwigia, Beiheft (Suppl.) 141: 263-274.), this species is generally found in brackish environments with a high-nutrient content. The entry of the species into freshwater can reflect the intermittent conditions of the river systems in northwest Ceará, where evaporation and low-levels of rainfall result in continuous rivers being converted into isolated pools, or dry up completely, during the dry season. High levels of evapotranspiration in the dry seasons lead to ion concentration and an increase in nutrients, especially in streams with no canopy cover ( Goméz et al. 2017 Goméz R, Arce MI, Baldwin DS & Dahm CN (2017) Water physicochemistry in intermittent rivers and ephemeral streams. In: Datry T, Bonada N & Boulton A (eds.) Intermittent rivers and ephemeral streams ecology and management. Academic Press, Cambridge. Pp. 109-134.; Olson 2019Olson JR (2019) Predicting combined effects of land use and climate change on river and stream salinity. Philosophical Transactions of the Royal Society, B: Biological Sciences 374: 20180005.). In addition, the pollution caused by domestic sewage exacerbates the accumulation of nutrients. The environmental characteristics of the Acaraú River in our study are very similar to those reported in records of C. marina by Cavalcante et al. ( 2013Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.) of a shallow urban river in Northeastern Brazil.
Finally, floristic studies are essential to understand the geographic distribution of taxa and to support future ecological studies. This study shows that the diatom diversity in Brazilian semi-arid region is underestimated, contributing to a more consistent understanding of diatom distribution in the country.
Acknowledgments
The authors thank the Center for Electron Microscopy of the Federal University of Paraná, for SEM availability; and to Eduardo Tusset, for operational assistance. Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES) provided a Master’s degree scholarship for MGRM; and TAVL was supported by a productivity grant from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (process number: 311876/2019-6). Authors are also grateful to Evelyn R. Nimmo, for edited the English language of the manuscript. To Prof. Elnatan B. de Souza, Prof. Maria Luiza R. C. Ribeiro and Prof. Lúcia Betânia S. Andrade from State University Vale do Acaraú, for their contribution in the fieldwork and laboratory equipments.
- Aké-Castillo JA, Okolodkov YB, Espinosa-Matías S, Merino-Virgilio FC, Herrera-Silveira JA & Ector L (2012) Cyclotella marina (Tanimura, Nagumo et Kato) Aké-Castillo, Okolodkov et Ector comb. et stat. nov. (Thalassiosiraceae): a bloom-forming diatom in the southeastern Gulf of Mexico. Nova Hedwigia, Beiheft (Suppl.) 141: 263-274.
- Albuquerque UP, Araújo EL, El-Deir ACA, Lima ALA, Souto A, Bezerra BM, Ferraz EMN, Freire EMX, Sampaio EVSB, Las-Casas FMG, Moura GJB, Pereira GA, Melo JG, Ramos MA, Rodal MJN, Schiel N, Lyra-Neves RM, Alves RRN, Azevedo-Júnior SM, Telino-Júnior WR & Severi W (2012) Caatinga revisited: ecology and conservation of an important Seasonal dry Forest. The Scientific World Journal 2012: 1-18.
- Álvarez-Blanco I, Blanco S, Cejudo-Figueiras C & Bécares E (2013) The Duero Diatom Index (DDI) for river water quality assessment in NW Spain: design and validation. Environmental Monitoring and Assessment 185: 969-981.
- Amorim CA, Dias AS & Ferreira RJ (2015) Microalgas perifíticas associadas à Nymphoides indica (L.) O. Kuntze em um reservatório do semiárido cearense. Caderno de Cultura e Ciência 14: 43-54.
- Andrade EM, Aquino DN, Chaves LCG & Lopes FB (2017) Water as capital and its uses in the caatinga. In: Silva JMC, Leal IR & Tabarelli M (eds.) Caatinga: the largest tropical dry forest region in South America. Springer International Publishing AG, New York. Pp. 281-302.
- Aquino EP, Oliveira ECC, Fernandes UL & Lacerda SR (2011) Phytoplankton in stabilization ponds in northeast Brazil. Brazilian Journal of Aquatic Sciences and Technology 15: 71-77.
- Bastos FH & Cordeiro AMN (2012) Fatores naturais na evolução das paisagens no semiárido brasileiro: Uma abordagem geral. Revista Geonorte 3: 464-476.
- Brassac NM, Atab DR, Landucci M, Visinoni ND & Ludwig TAV (1999) Diatomáceas cêntricas de rios da região de abrangência da usina hidrelétrica de Salto Caxias, PR (Bacia do Rio Iguaçu) Acta Botanica Brasilica 13: 277-289.
- Buczkó K, Ognjanova-Rumenova N & Magyari E (2010) Taxonomy, morphology and distribution of some Aulacoseira taxa in Glacial Lakes in the south Carpathian region. Polish Botanical Journal 55: 149-163.
- Cavalcante KP, Tremarin PI & Ludwig TAV (2013) Taxonomic studies of centric diatoms (Diatomeae): unusual nanoplanktonic forms and new records for Brazil. Acta Botanica Brasilica 27: 237-251.
- Chung MH, Yoon WD & Lee JB (2010) Morphological description of Cyclotella atomus var. marina (Bacillariophyceae): newly reported in Korean waters Algae 25: 57-64.
- Compère P (1982) Taxonomic revision of the diatom genus Pleurosira (Eupodiscaceae). Bacillaria 5: 165-190.
- Costa ARS, Amorim CA, Nascimento KJ, Dias AS, Ferreira RJ & Lacerda SR (2015) Caracterization of microalgae periphytic community in a reservoir semiarid cearense. Caderno de Cultura e Ciência 14: 43-59.
- Cox EJ (2013) Recognising and defining taxonomic boundaries in ‘well-known’ freshwater diatoms and its relevance to water quality evaluation. Diatomededelingen 37: 17-26.
- Cox EJ (2014) Diatom identification in the face of changing species concepts and evidence of phenotypic plasticity. Journal of Micropalaeontology 33: 111-120.
- Crawford RM, Likhoshway YV & Jahn R (2003) Morphology and identity of Aulacoseira Italica and typification of Aulacoseira (Bacillariophyta). Diatom Research 18: 1-19.
- Dajoz R (2005) Princípios de ecologia. 7ª ed. Artmed, Porto Alegre. 520p.
- Da Silva WJ, Nogueira IS & Souza MGM (2011) Catálogo de diatomáceas da região Centro-Oeste brasileira. Iheringia Série Botânica 66: 61-86.
- Dalu T & Froneman PW (2016) Diatom-based water quality monitoring in southern Africa: challenges and future prospects. Water SA 42: 551-559.
- Ehrenberg CG (1843) Verbreitung und Einfluss des mikroskopischen Lebens in Süd- und Nord-Amerika. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1841: 291-445.
- Eskinazi-Leça E, Moura CWN, Cunha MGGS, Santiago MF, Borges GCP, Lima JC, Silva MH, Ferreira LC, Aquino E, Da Silva WJ & Menezes M (2015) Bacillariophyceae. In: Lista de espécies da flora do Brasil. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Available at < http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB119495>. Access on 28 November 2019.
» http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB119495 - Faustino SB, Fontana L, Bartozek ECR, Bicudo CEM & Bicudo DC (2016) Composition and distribution of diatom assemblages from core and surface sediments of a water supply reservoir in Southeastern Brazil. Biota Neotropica 16: e20150129.
- Ferrari F & Ludwig TAV (2007) Coscinodiscophyceae, Fragilariophyceae e Bacillariophyceae (Achnanthales) dos rios Ivaí, São João e dos Patos, bacia hidrográfica do Rio Ivaí, município de Prudentópolis, PR, Brasil. Acta Botanica Brasilica 21: 421-441.
- Gargas CB, Theriot EC, Ashworthb MP & Johansen JR (2018) Phylogenetic analysis reveals that the ‘radial centric’ diatom Orthoseira Thwaites (Orthoseiraceae, Bacillariophyta) is a member of a ‘multipolar’ diatom lineage. Protist 169: 803-825.
- Genkal SI & Bilous OP (2015) Centric Diatoms (Centrophyceae) of the lower portion of the Southern Bug river (Ukraine). International Journal on Algae 17: 339-350.
- Genkal SI & Yarmoshenko LP (2013) Centric Diatoms (Bacillariophyta) of the cooling pond of the Khmelnitskiy Nuclear Power Station (Ukraine). Hydrobiological Journal 49: 51-63.
- Genkal SI & Okhapkin AG (2013) Centric Diatoms (Centrophyceae) of the lower reaches of the Oka river (Russian Federation). Hydrobiological Journal 49: 41-57.
- Genkal SI (2014) Morphology, taxonomy, ecology and distribution of Cyclotella meduanae Germain (Bacillariophyta). Nova Hedwigia Beiheft 143: 127-140.
- Genkal SI, Bondarenko NA & Popovskaya GI (2007) New representative of the genus Discostella Houck et Klee (Bacillariophyta) from the Eastern Baikal area (Russia). International Journal on Algae 9: 359-364.
- Germain H (1981) Flore des diatomées Diatomophycées eaux douces et saumâtres du Massif Armoricain et des contrées voisines d’Europe occidentale. Collection "Faunes et Flores Actuelles". Société Nouvelle des Editions Boubée, Paris. 444p.
- Goméz R, Arce MI, Baldwin DS & Dahm CN (2017) Water physicochemistry in intermittent rivers and ephemeral streams. In: Datry T, Bonada N & Boulton A (eds.) Intermittent rivers and ephemeral streams ecology and management. Academic Press, Cambridge. Pp. 109-134.
- Guerrero JM & Echenique RO (2006) Discostella taxa (Bacillariophyta) from the Río Limay basin (northwestern Patagonia, Argentina). European Journal of Phycology 41: 83-96.
- Hasle GR (1983) The marine planktonic diatoms Thalassiosira oceanica sp. nov. and T. partheneia. Journal of Phycology 19: 220-229.
- Hendey NI (1972) Some marine diatoms from the Galapagos Islands. Nova Hedwigia 22: 371-422.
- Hevia-Orube J, Orive E, David H, Díez A, Laza-Martínez A, Miguel I & Seoane S (2015) Molecular and morphological analyses of solitary forms of brackish Thalassiosiroid diatoms (Coscinodiscophyceae), with emphasis on their phenotypic plasticity. European Journal of Phycology 51: 1-20.
- Houk V & Klee R (2004) The stelligeroid taxa of the genus Cyclotella (Kutz.) Brébisson (Bacillariophyceae) and their transfer to the new genus Discostella gen. nov. Diatom Research 19: 203-228.
- Houk V (1993) Some morphotypes in the Orthoseira roeseana complex. Diatom Research 8: 385-402.
- Houk V (2003) Atlas of freshwater centric diatoms with a brief key and descriptions. Part I. Melosiraceae, Orthoseiraceae, Paraliaceae and Aulacoseiraceae. In: Poulícková A (ed.) Czech Phycology Supplement, Olomouc. 111p.
- Houk V, Klee R & Tanaka H (2010) Atlas of freshwater centric diatoms with a brief key and descriptions. Part III. Stephanodiscaceae A. Cyclotella, Tertiarius, Discostella. Fottea 10: 1-498.
- Huber-Pestalozzi G (1942) Diatomeen. In: Thienimann A (ed.) Das phytoplankton des susswassers, systematik und biologie. Stuttgart E. Schweizerbartsche Verlagsbuchhandlung 2: 367-549.
- Hustedt F (1937) Systematische und ökologische Untersuchungen über die Diatomeen-Flora von Java, Bali und Sumatra nach dem Material der Deutschen Limnologischen Sunda-Expedition. Archiv für Hydrobiologie (Supplement) 15: 131-177.
- Hustedt F (1942) Süßwasser-Diatomeen des indomalayischen Archipels und der Hawaii-Inseln. Nach dem Material der Wallacea-Expedition. Internationale Revue der gesamten Hydrobiologie und Hydrographie 42: 1-252.
- IPECE (2008) características geográficas, recursos naturais e meio ambiente. Ceará em números. Instituto de Pesquisa e Estratégia Econômica do Ceará. Pp. 30-31. Available at < http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2008/completa/>. Access on 28 November 2019.
» http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2008/completa/ - IPECE (2016) Características geográficas, recursos naturais e meio ambiente. Ceará em números. Instituto de Pesquisa e Estratégia Econômica do Ceará. Pp. 21-44. Available at < http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2016/completa/>. Access on 25 November 2019.
» http://www2.ipece.ce.gov.br/publicacoes/ceara_em_numeros/2016/completa/ - Jiménez JA, Magos YB & Camarillo VHS (2017) Morphological and environmental characterization of Terpsinoë musica (Biddulphiaceae, Bacillariophyceae) in tropical streams from Mexico. Diatom Research 32: 185-193.
- Joh G (2010) Algal flora of Korea. Chrysophyta, Bacillariophyceae: centrales. Freshwater diatoms I. National Institute of Biological Resources 3: 1-161.
- Julius ML & Theriot EC (2010) The diatoms: a primer. In: Smol JP & Stoermer EF (eds.) The diatoms: applications for the environmental and earth sciences. 2nd ed. Cambridge University Press, Cambridge. Pp. 8-22.
- Karthick B & Kociolek JP (2011) Four new centric diatoms (Bacillariophyceae) from the Western Ghats, South India. Phytotaxa 22: 25-40.
- Kelly MG, Cazaubon A, Coring E, Dell’uomo A, Ector L, Goldsmith B, Guasch H, Hürlimann J, Jarlman A, Kawecka B, Kwandrans J, Laugaste R, Lindstrøm EA, Leitao M, Marvan P, Padisák J, Pipp E, Prygiel J, Rott E, Sabater S, Van-Dam H & Vizinet J (1998) Recommendations for the routine sampling of diatoms for water quality assessments in Europe. Journal of Applied Phycology 10: 215-224.
- Kiss KT, Klee R, Ector L & Ács É (2012) Centric diatoms of large rivers and tributaries in Hungary: morphology and biogeographic distribution. Acta Botanica Croatica 71: 311-363.
- Krammer K (1991) Morphology and taxonomy in some taxa of the genus Aulacoseira Thwaites (Bacillariophyceae). II. Taxa in the A. granulata-, italica- and lirata-groups. Nova Hedwigia 53: 477-496.
- Kützing FT (1844) Die Kieselschaligen Bacillarien oder Diatomeen. Zu finden bei W. Köhne, Nordhausen. Pp. [i-vii], [1]-152, pls 1-30.
- Lai GG, Padedda BM, Virdis T, Sechi N & Lugliè A (2014) Benthic diatoms as indicators of biological quality and physical disturbance in Mediterranean watercourses: a case study of the Rio Mannu di Porto Torres basin, northwestern Sardinia, Italy. Diatom Research 29: 11-26.
- Landucci M & Ludwig TAV (2005) Diatomáceas de rios da Bacia Hidrográfica Litorânea, PR, Brasil: Coscinodiscophyceae e Fragilariophyceae. Acta Botanica Brasilica 19: 345-357.
- Leal IR, Silva JMC & Barros MLB (2003) Ecologia e conservação da caatinga. Editora Universitária da UFPE, Recife. 822p.
- Leal IR, Silva JMC, Tabarelli M & Lacher T (2005) Changing the course of biodiversity conservation in the Caatinga of Northeastern Brazil. Conservation Biology 19: 701-706.
- Leuduger-Fortmorel G (1898) Diatomées marines da costa ocidental de África. Saint-Brieuc Francisque Guyon, Saint-Brieuc. 41p.
- Lobo EA, Schuch M, Heinrich CG, Costa AB, Düpont A, Wetzel CE & Ector L (2015) Development of the Trophic Water Quality Index (TWQI) for subtropical temperate Brazilian lotic systems. Environmental Monitoring and Assessment 187: 1-13.
- Lobo EA, Wetzel CE, Ector L, Katoh K, Blanco S & Mayama S (2010) Response of epilithic diatom communities to environmental gradients in subtropical temperate Brazilian rivers. Limnetica 29: 10-20.
- Ludwig TAV & Tremarin PI (2017) Bacillariophyta. In: Bicudo CEM & Menezes M (eds.) Gênero de algas de águas continentais do Brasil: chave para identificação e descrição. 3ª ed. RiMa, Rio de Janeiro. Pp. 435-503.
- Ludwig TAV, Flores TL, Moreira-Filho H & Veiga LAS (2004) Inventário florístico das diatomáceas (Ochrophyta) de lagoas do Sistema Hidrológico do Taim, Rio Grande do Sul, Brasil: Coscinodiscophyceae. Iheringia, Série Botânica 59: 97-106.
- Luttenton MR, Pfiester LA & Timpano P (1986) Morphology and growth habitat of Terpsinoe musica Ehr. (Bacillariophyceae). Castanea, Journal of the Southern Appalachian Botanical Club. Morgantown 51: 175-182.
- Medlin LK & Kaczmarska I (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Journal Phycologia 43: 245-270.
- Menezes M, Bicudo CEM & Moura CWN (2015) Update of the Brazilian floristic list of algae and cyanobacteria. Rodriguésia 66: 1047-1062.
- Metzeltin D & Lange-Bertalot H (2007) Tropical Diatoms of South America II. Special remarks on biogeographic disjunction. Iconographia Diatomologica (H. Lange-Bertalot, ed.). Vol. 18. Koeltz Scientifc Books, Stuttgart. 877p.
- Metzeltin D, Lange-Bertalot H & García-Rodríguez F (2005) Diatoms of Uruguay, compared with other taxa from South America and elsewhere. Iconographia Diatomologica (H. Lange-Bertalot, ed.). Vol. 15. Koeltz Scientifc Books, Stuttgart. 736p.
- Morandi LL, Ritter LMO, Moro RS & Bicudo CEM (2006) Criptógamos do Parque Estadual do Ipiranga, São Paulo, SP. Algas, 20: Coscinodiscophyceae. Hoehnea 33: 115-122.
- Moreira-Filho H & Valente-Moreira IM (1981) Avaliação taxonômica e ecológica das diatomáceas (Bacillariophyceae) epífitas em algas pluricelulares obtidas nos litorais dos estados dos estados do Paraná, Santa Catarina e São Paulo. Boletim Museu Botânico Municipal de Curitiba 47: 1-17.
- Moro MF, Macedo MB, Moura-Fé MM, Castro ASF & Costa RC (2015) Vegetation, phytoecological regions and landscape diversity in Ceará state, northeastern Brazil. Rodriguésia 66: 717-743.
- Nardelli MS, Bueno NC, Ludwig TAV, Tremarin PI & Bartozek ECR (2014) Coscinodiscophyceae and Fragilariophyceae (Diatomeae) in the Iguaçu river, Paraná, Brazil. Acta Botanica Brasilica 28: 127-140.
- Nascimento F (2011) Categorização de usos múltiplos dos recursos hídricos e problemas ambientais. Revista da Associação Nacional de Pós-graduação e Pesquisa em Geografia 7: 81-97.
- Öberg H, Risberg J & Stabell B (2009) Morphology, valve ultrastructure and stratigraphical variability of Discostella taxa in a tropical crater lake, northern Tanzania. Diatom Research 24: 341-356.
- Olson JR (2019) Predicting combined effects of land use and climate change on river and stream salinity. Philosophical Transactions of the Royal Society, B: Biological Sciences 374: 20180005.
- Patrick R (1940a) Diatoms of Northeastern Brazil. Part I. Coscinodiscaceae, Fragilariaceae and Eunotiaceae. Proceedings of the Academy of Natural Sciences of Philadelphia 92: 191-226.
- Patrick R (1940b) Some new diatoms from Brazil. Notulae Naturae of the Academy of Natural Sciences of Philadelphia 59: 1-7.
- Pfitzer E (1871) Untersuchungen über bau und entwicklung der bacillariaceen (Diatomaceen). In: von Hanstein JLER (ed.) Botanische abhandlungen aus dem Gebiet der Morphologie und Physiologie. Vol. 1. A. Marcus, Bonn. 189p.
- Reimann BEF, Lewin JMC & Guillard RRL (1963) Cyclotella cryptica, a new brackish-water diatom species. Phycologia 3: 75-84.
- Round FE, Crawford RM & Mann DG (1990) The diatoms: biology and morphology of the genera. Cambridge University Press, Cambridge. 747p.
- Sala S & Ramírez JJ (2008) Cyclotella katiana sp. nov. from La Reina Swamp, Parque Nacional Natural los Katíos, Colombia. Diatom Research 23: 147-167.
- Santos JC, Leal IR, Almeida-Cortez JS, Fernandes GW & Tabarelli M (2011) Caatinga: the scientific negligence experienced by a dry tropical forest. Tropical Conservation Science 4: 276-286.
- Schmidt A (1812-1899) Verzeichniss der. In: A. Schmidt's Atlas der Diatomaceenkunde: Heft 1-36 (Serie I-III) abgebildeten Arten und benannten Varietäten nebst den mit angeführten Synonymen. Topics Diatoms. O.R. Reisland, Leipzig. Pp. 350.
- Schultz ME (1971) Salinity-related polymorphism in the brackish-water diatom Cyclotella cryptica. Canadian Journal of Botany 49: 1285-1289.
- Silva JMC, Barbosa LCF, Leal IR & Tabarelli M (2017) The Caatinga: understanding the challenges. In: Silva JMC, Leal IR & Tabarell M (eds.) Caatinga: The largest tropical dry forest region in South America. Springer International Publishing AG, Cham. Pp. 3-19.
- Silva-Lehmkuhl AM, Tremarin PI, Vercellino IS & Ludwig TAV (2019) Periphytic diatoms from an oligotrophic lentic system, Piraquara I reservoir, Paraná state, Brazil. Biota Neotropica 19: e20180568.
- Simonsen R (1974) The diatom plankton of the Indian Ocean Expedition of R/V “Meteor”, 1964-65. “Meteor” Forschungsergbnisse, Reih D-Biol 19: 1-66.
- Simonsen R (1979) The diatom system: ideas on phylogeny. Bacillaria 2: 9-71.
- Spaulding SA & Kociolek JP (1998) The diatom genus Orthoseira: ultrastructure and morphological variation in two species from Madagascar with comments on nomenclature in the Genus. Diatom Research 13: 133-147.
- Stachura-Suchoples K & Williams DM (2009) Description of Conticribra tricircularis, a new genus and species of Thalassiosirales, with a discussion on its relationship to other continuous cribra species of Thalassiosira Cleve (Bacillariophyta) and its freshwater origin. European Journal of Phycology 44: 477-486.
- Stevenson J (2014) Ecological assessments with algae: a review and synthesis. Journal Phycology 50: 437-461.
- Syvertsen EE & Hasle GR (1984) Thalassiosira bulbosa Syvertsen sp. nov., an Artic marine diatom. Polar Biology 3: 167-172.
- Tanaka H (2007) Taxonomic studies of the genera Cyclotella (Kützing) Brébisson, Discostella Houk et Klee and Puncticulata Håkansson in the Family Stephanodiscaceae Glezer et Makarova (Bacillariophyta) in Japan. Bibliotheca Diatomologica, Band 53. J Cramer, Berlim. 204p.
- Tanimura Y, Nagumo T & Kato M (2004) A new variety of Cyclotella atomus from Tokyo Bay, Japan, C. atomus var. marina var. nov. Bulletin of the National Science Museum Series C (Geology and Paleontology) 30: 5-11.
- Tremarin PI, Ludwig TV & Torgan LC (2013) Morphological variation and distribution of the freshwater diatom Aulacoseira ambigua (Grunow) Simonsen in Brazilian continental environments. Iheringia Série Botânica 68: 139-157.
- Tuji A & Williams DM (2006) Type examination of Cyclotella wortereckii Hust. (Bacillariophyceae) with special attention to the position of its rimoportula. Bulletin of the National Science Museum Series B (Botany) 32: 15-17.
- Tuji A (2018) A new freshwater diatom, Terpsinoë muninensis sp. nov., from the Ogasawara Islands, Japan. Memoirs of the National Science Museum (Tokyo) 52: 5-15.
- Vieira RDS, Lacerda SR, Oliveira ECC, Nascimento KJ & Dias AS (2013) Biodiversidade de microalgas perifíticas do Rio da Batateira (Sítio Fundão, Crato, CE). Cadernos de Cultura e Ciência 12: 7-15.
- Virta L, Gammal J, Järnström M, Bernard G, Soininen J, Norkko J & Norkko A (2019) The diversity of benthic diatoms affects ecosystem productivity in heterogeneous coastal environments. Ecology 100: 1-11.
- Wu S (2013) Terpsinoë musica. Diatoms of North America. Available at < https://diatoms.org/species/terpsinoe_musica>. Access on 23 June 2020.
» https://diatoms.org/species/terpsinoe_musica
Edited by
Area Editor:
Data availability
Data citations
Eskinazi-Leça E, Moura CWN, Cunha MGGS, Santiago MF, Borges GCP, Lima JC, Silva MH, Ferreira LC, Aquino E, Da Silva WJ & Menezes M (2015) Bacillariophyceae. In: Lista de espécies da flora do Brasil. Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Available at < http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB119495>. Access on 28 November 2019.
Publication Dates
-
Publication in this collection
21 Nov 2022 -
Date of issue
2022
History
-
Received
25 Feb 2021 -
Accepted
29 Oct 2021