Abstract
Background
Toxic cyanobacterial blooms are recurrent in Patos Lagoon, in southern Brazil. Among cyanotoxins, [D-Leu1] microcystin-LR is the predominant variant whose natural cycle involves water and sediment compartments. This study aimed to identify and isolate from sediment a bacterial strain capable of growing on [D-Leu1] microcystin-LR. Sediment and water samples were collected at two distinct aquatic spots: close to the Oceanographic Museum (P1), in Rio Grande City, and on São Lourenço Beach (P2), in São Lourenço do Sul City, southern Brazil.
Methods
[D-Leu1] microcystin-LR was isolated and purified from batch cultures of Microcystis aeruginosastrain RST9501. Samples of water and sediment from Rio Grande and São Lourenço do Sul were collected. Bacteria from the samples were allowed to grow in flasks containing solely [D-Leu1] microcystin-LR. This strain named DMSX was isolated on agar MSM with 8 g L−1 glucose and further purified on a cyanotoxin basis growth. Microcystin concentration was obtained by using the ELISA immunoassay for microcystins whereas bacterial count was performed by epifluorescence microscopy. The genus Pseudomonas was identified by DNA techniques.
Results
Although several bacterial strains were isolated from the samples, only one, DMXS, was capable of growing on [D-Leu1] microcystin-LR. The phylogenetic analysis of the 16S rRNA gene from DMXS strain classified the organism as Pseudomonas aeruginosa. DMXS strain incubated with [D-Leu1] microcystin-LR lowered the amount of toxin from 1 μg.L−1 to < 0.05 μg.L−1. Besides, an increase in the bacterial count–from 71 × 105 bacteria.mL−1 to 117 × 105 bacteria.mL−1–was observed along the incubation.
Conclusions
The use of bacteria isolated from sediment for technological applications to remove toxic compounds is viable. Studies have shown that sediment plays an important role as a source of bacteria capable of degrading cyanobacterial toxins. This is the first Brazilian report on a bacterium–of the genus Pseudomonas–that can degrade [D-Leu1] microcystin-LR, the most frequent microcystin variant in Brazilian freshwaters.
Biodegradation; Microcystin; Sediment; Patos Lagoon; Estuary; Brazil
Background
Cyanobacteria are also known as blue-green algae, because they have characteristics of both algae and bacteria, although they are now classified as bacteria. These organisms are capable of producing toxins, called cyanotoxins, and their blue-green color comes from the pigments which gives their ability to photosynthesize. In waterbodies used for water supply, cyanobacterial blooms can pose serious threats to animals and humans.
Toxic cyanobacterial blooms have been reported worldwide [11 Kotak BG, Kenefick SL, Fritz DL, Rousseaux CG, Prepas EE, Hrudey SE: Occurrence and toxicological evaluation of cyanobacterial toxins in Alberta lakes and farm dugouts. Water Res 1993, 27(3):495-506.]-[77 Matthiensen A, Beattie KA, Yunes JS, Kaya K, Codd GA: [D-Leu1] Microcystin-LR, from the cyanobacterium Microcystis RST 9501 and from a Microcystis bloom in the Patos Lagoon estuary, Brazil. Phytochemistry 2000, 55(5):383-387.]. A serious case of cyanobacterial contamination occurred in Brazil. A hemodialysis clinic in the city of Caruaru, in northeastern Pernambuco, used water contaminated with cyanotoxins, which led to more than 60 patient deaths [88 Jochimsen EM, Carmichael WW, An J, Cardo DM, Cookson ST, Holmes CEM, et al.: Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N Engl J Med 1998, 338:873-878.],[99 Pouria S, De Andrade A, Barbosa J, Cavalcanti RL, Barreto VT, Ward CJ, et al.: Fatal microcystin intoxication in haemodialysis unit in Caruaru. Brazil Lancet 1998, 352(9121):21-26.].
In Patos Lagoon, in the southernmost part of Brazil, toxic cyanobacterial blooms have been observed for 18 years [44 Yunes JS, Salomon PS, Matthiensen A, Beattie KA, Raggett SL, Codd GA: Toxic blooms of cyanobacteria in the Patos Lagoon Estuary, southern Brazil. J Aquat Ecosystem Health 1996, 5(4):223-229.],[1010 Yunes JS, Niencheski LF, Salomon P, Parise M, Beattie KA, Raggett SL, et al.: Effect of nutrient balance and physical factors on blooms of toxic cyanobacteria in the Patos Lagoon, southern Brazil. Verh Inter Verein Limnol 1998, 26:1796-1800.] as seen in Figure 1.
The Patos Lagoon is a large waterbody that covers an area of 10,360 km2. Since several freshwater rivers run into it, it involves an even higher surface area of 200,000 km2 in the Rio Grande do Sul state. The estuarine area comprises 10% of the lagoon and is fundamental to fish species, crustaceans and mollusks, supplying food, shelter and reproduction sites (several species come to the estuary to spawn). The economic activities of the area lies on fishery, waterways and recreational places [44 Yunes JS, Salomon PS, Matthiensen A, Beattie KA, Raggett SL, Codd GA: Toxic blooms of cyanobacteria in the Patos Lagoon Estuary, southern Brazil. J Aquat Ecosystem Health 1996, 5(4):223-229.],[1111 Lemes GA, Kersanach R, Pinto L da S, Dellagostin OA, Yunes JS, Matthiensen A:Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol Environ Saf 2008, 69(3):358-365.]. Therefore, toxic cyanobacterial blooms affect not only estuarine organisms, but also the local economy.
Cyanobacterial blooms in Patos Lagoon are extremely toxic since mouse bioassays detected lethal concentrations below 100 μg.kg−1 of body weight [1212 Matthiensen A, Yunes JS, Codd GA: Ocorrência, distribuição e toxicidade de cianobactérias no estuário da Lagoa dos Patos. RS Rev Bras Biol 1999, 59(3):361-376.]. These blooms are associated with pH (7.5-8.0), temperatures (>20°C) and high concentration of nutrients [44 Yunes JS, Salomon PS, Matthiensen A, Beattie KA, Raggett SL, Codd GA: Toxic blooms of cyanobacteria in the Patos Lagoon Estuary, southern Brazil. J Aquat Ecosystem Health 1996, 5(4):223-229.]. Among the toxins present in cyanobacterial blooms are microcystins and cyclic heptapeptides that have more than 65 structural variants with variable toxicity. Microcystins may trigger tumor development and have been associated with acute and chronic health problems in humans and other animals.
Microcystis aeruginosa is the main toxic species found in Patos
Lagoon and is known for producing [D-Leu1] microcystin-LR [77 Matthiensen A, Beattie KA, Yunes JS, Kaya K, Codd GA: [D-Leu1]
Microcystin-LR, from the cyanobacterium Microcystis RST 9501
and from a Microcystis bloom in the Patos Lagoon estuary, Brazil.
Phytochemistry 2000,
55(5):383-387.],[1313 Matthiensen A. Environmental and laboratory studies on the
properties and fate of microcystins from the cyanobacterium Microcystis. PhD
thesis. University of Dundee, UK; 2000:307. [available at University of Dundee
Library: http://www.dundee.ac.uk/library/search/].
http://www.dundee.ac.uk/library/search/...
].
This variant comprises 90% of the total amount of toxins produced intracellularly
and is one of the toxins with the highest toxicity [1313 Matthiensen A. Environmental and laboratory studies on the
properties and fate of microcystins from the cyanobacterium Microcystis. PhD
thesis. University of Dundee, UK; 2000:307. [available at University of Dundee
Library: http://www.dundee.ac.uk/library/search/].
http://www.dundee.ac.uk/library/search/...
]-[1515 Sivonen K, Jones G: Cyanobacterial toxins. In
Toxic cyanobacteria in water: A guide to their public health
consequences, monitoring and management. World Health Organization,
E&FN Spon, London; 1999:41-111.]. Investigation on the
biodegradation of cyanobacterial toxins have been carried out in laboratories and in
fieldwork aiming at avoiding damage caused by these blooms [1616 Cousins IT, Bealing DJ, James HA, Sutton A: Biodegradation of
microcystin-LR by indigenous mixed bacterial populations.
Water Res 1996,
30(2):481-485.]-[1919 Ji RP, Lu XW, Li XN, Pu YP: Biological degradation of algae
and microcystins by microbial enrichment on artificial media.
Ecol Eng 2009,
35(11):1584-1588.].
In the south of Brazil, the first study on the biodegradation of [D-Leu1] microcystin-LR was performed with samples of Patos Lagoon water [2020 Matthiensen A, Metcalf JS, Ferreira AHF, Yunes JS, Codd GA: Biodegradation and biotransformation of microcystin by aquatic microbes in estuarine waters from the Patos Lagoon, RS, Brazil. In Mycotoxins and Phycotoxins in perpective at the turn of the Millenium. Edited by Koe WJ, Samson SA, Egmond HP, Gilbert J, Wageningen SM. Posen & Looyen, Brazil; 2000:527-36.]. Afterwards, another research was conducted to identify a specific strain that is capable of degrading microcystin-LR and [D-Leu1] microcystin-LR originated from the estuary of Patos Lagoon [1111 Lemes GA, Kersanach R, Pinto L da S, Dellagostin OA, Yunes JS, Matthiensen A:Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol Environ Saf 2008, 69(3):358-365.].
Since several studies recommend the biodegradation of cyanobacterial toxins as a safe and viable manner to remove toxic compounds from water, the current work aimed to isolate and identify a bacterial strain collected from the sediment of Patos Lagoon which is able to degrade the toxin [D-Leu1] microcystin-LR [1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.],[2121 Ishii H, Nishijima M, Abe T: Characterization of degradation process of cyanobacterial hepatotoxins by a gram-negative aerobic bacterium. Water Res 2004, 38(11):2667-2676.],[2222 Tsuji K, Asakawa M, Anzai Y, Sumino T, Harada KI: Degradation of microcystins using immobilized microorganism isolated in an eutrophic lake. Chemosphere 2006, 65(1):117-124.].
Methods
Cultures
Experiments were carried out with the toxin extracted from cultures of the cyanobacteriumMicrocystis aeruginosa RST9501. The strain was isolated from water collected in Patos Lagoon and deposited in the Culture Collection of the Laboratory of Cyanobacteria and Phycotoxins. Cultures were maintained in 1 L Erlenmeyer flasks with BG-11 medium [2323 Stanier RY, Kunisawa R, Mandel M, Cohen-Bazire G: Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol Rev 1971, 35(2):171-205.]. Culture flasks were kept in FANEM 347 growth chambers at 20°C ± 2°C in light–dark cycles of 12 hours. The cell growth was performed in order to yield a major variant of microcystin, [D-Leu1] microcystin-LR, for use in the experiments.
Extraction and purification of microcystin
The toxin [D-Leu1] microcystin-LR was extracted from cells according to Lawton et al.[2424 Lawton LA, Edwards C, Codd GA: Extraction and High-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst (Cambridge, U K) 1994, 119:1525-1530.]. Cultures were centrifuged (Hermle Z323, Labnet, Germany) at 8,000 g for 20 minutes. Afterwards, the centrifuged material was frozen and lyophilized (Edwards MicroModulyo, UK). Then, 250 mL of acetic acid and distilled water (5% v/v) was added for each gram of lyophilized material. The mixture was agitated for an hour and again centrifuged at 8,000 g for ten minutes. The supernatant (S1) was stored. The resulting material (pellet) of this centrifugation went through a second centrifugation process that was similar to the first one. After the centrifugation at 8,000 g for ten minutes, the pellet was discharged and the second supernatant (S2) was added to the previous (S1). Both resulting supernatants (S1 + S2 = 500 mL) were stored in a freezer for 24 hours. After thawing, they were centrifuged at 8,000 g for ten minutes and the resulting pellet was discharged.
In the second step, total resultant supernatants were filtered through a peristaltic pump (Millipore Corporation, USA) which was coupled to a Sep-Pak 3 mL/1 g C18 cartridge. The cartridge was previously activated by addition of 10 mL of methanol, followed by 10 mL of distilled water. Afterwards, the supernatants were filtered through the cartridges; these containers were frozen for 24 hours. After thawing, the cartridges were eluted with 20 mL of methanol (100%). The extracts that resulted from the passage of methanol through the cartridges were dried in a rotary evaporator with a vacuum system at 40°C. After evaporation, residues were resuspended in 500 μL of methanol (100%) twice and were then analyzed by high performance liquid chromatography (HPLC–Shimadzu SCL-10Avp, Japan) to determine the concentration of microcystins. The resulting toxin (Figure 2) was used in the biodegradation experiments.
Water and sediment collection
In Patos estuary, water and sediment samples were collected at spot P1, near the Oceanographic Museum (32.0246S; 52.1062 W) that belongs to the Federal University of Rio Grande (FURG). This spot was chosen because bacteria able to degrade cyanotoxins had already been found in that environment [1111 Lemes GA, Kersanach R, Pinto L da S, Dellagostin OA, Yunes JS, Matthiensen A:Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol Environ Saf 2008, 69(3):358-365.]. In São Lourenço do Sul city (31.3690 S; 51.9620 W), water samples were collected on the beach, at spot P2 (Figure 3), a place where heavy Microcystis blooms were seen, so they also could be the source of heterotrophic bacteria.
Surface water samples were collected in a 1-L plastic bucket and stored in clean plastic bottles. Temperature (Incoterm thermometer, Brazil) and salinity (Quimis salinometer, Brazil) were measured at the site whereas the pH was measured in the laboratory (Digimed pH meter, Brazil).
Sediment was only collected at P1, near the Oceanographic Museum, about 500 g with PVC pipes (75 cm in diameter) stored in clean plastic bottles and taken to the laboratory.
Afterwards, water and sediment samples were stored in styrofoam boxes and taken to the laboratory, where they were disposed in 250 mL minimal salt media (MSM) and vortexed for 30 minutes. All samples were collected and analyzed in triplicate.
Isolation of bacterial strains
To isolate bacterial strains, water samples were filtered through Whatman filters (7.0 cm, pore 0.45 μm), and, then, spread on Petri dishes with MSM and addition of 8 g.L−1 glucose anhydrous and 15 g L−1 agar [2525 Morsen A, Rehm HJ: Degradation of phenol by mixed culture of Pseudomonas putida and Crytococcus elinonii adsorbed on activated carbon. Appl Microbiol Biotechnol 1987, 26:283-288.]. Sediment samples were centrifuged (Hermle Z323, Labnet, Germany) at 12,000 rpm for ten minutes and the supernatant was filtered in Whatman cellulose acetate membrane filters (7.0 cm, pore 0.45 μm). The remaining pellet was discharged. Several strains were isolated from the water collected in both spots and from the sediment collected at P2, strains were catalogued and kept in cultures for the biodegradation tests (Table 1).
Bacteria isolated from water samples collected at two spots in Patos Lagoon: P1, in the Patos Lagoon estuary (near the Oceanographic Museum) and P2, on São Lourenço Beach (Rio Grande do Sul state, Brazil)
Biodegradation experiment of [D-Leu1] microcystin-LR
The biodegradation experiment of [D-Leu1] Microcystin–LR was carried out in a first screening basis, with each strain that was isolated from water and sediment samples collected at spots P1 and P2.
The experiment was conducted with 30 mL MSM in each flask, which had previously been autoclaved at 120 kgf/cm2 for 30 minutes. Three amber flasks were kept for the treatment and three were kept for control. Eight microliters of [D-Leu1] microcystin-LR at 3.7 μg.L−1 was added to each flask, so that the approximate concentration was 1 μg.L−1.
Treatment flasks had bacterial samples from the culture dishes whereas control flasks did not have bacterial strains.
The experiment was kept in a microbiological heater Certomat® BS-1 (Germany) at 27°C, in dim light to prevent the toxin from degrading due to intense light.
During the experiment, samples were collected for analysis every three days, on average; all handling was done in a microbiological chamber with a Bunsen burner. Total sampling time was 30 days. Medium samples were labeled and stored in a freezer at –20°C for further analysis by ELISA immunoassay.
Selection and isolation of the strain to perform the biodegradation assay
At the end of the experiment with sediment, when the concentration of the toxin [D-Leu1] microcystin-LR reached “zero”, a sample of this bacterial strain was removed from the flasks and inoculated in MSM on a Petri dish with the addition of 8 g.L−1 glucose anhydrous and 15 g.L−1 agar. The strain was then denominated DMXS. In order to guarantee the purity of the isolated, the strain was inoculated in Petri plates containing growth media with pure D-Leu microcystin-LR. A pure isolate was considered a colony (single form) which was repeatedly replicated successfully.
Evaluation of toxin [D-Leu1] microcystin-LR concentration by Elisa Immunoassay
Assessment of toxin levels was carried out by ELISA immunoassay with specific antibodies for microcystins. This test has high sensitivity and detects toxins in concentrations as low as 0.05 up to 2.5 μg.L−1. The ELISA kit (Abraxis, USA) was used in accordance with the methodology recommended by the manufacturer.
Bacterial count
Bacterial count was performed through epifluorescence microscopy in a Zeiss Axioplan microscope (Germany) and using the software UTHSCSA Image Tool (Version 3.0) (http://compdent.uthscsa.edu/dig/download.html webcite). Samples were filtered through polycarbonate membranes (Nuclepore, Whatman, UK; pore 0.2 μm) and colored with Irgalan black. Samples were then dyed with acridine orange in accordance with the methodology proposed by Hobbie et al.[2626 Hobbie JE, Daley RJ, Jasper S: Use of nuclepore filters for counting bacteria by fluorescence microscopy. Appl Environ Microbiol 1977, 33(5):1225-1228.] to count bacteria in samples fixed with Lugol’s solution [2727 Pomroy AJ: Direct counting of bacteria preserved with Lugol Iodine solution. Appl Environ Microbiol 1984, 47(5):1191-1192.],[2828 Nishino SF: Direct acridine orange counting of bacteria preserved with acidified Lugol Iodine. Appl Environ Microbiol 1986, 52(3):602-604.]. This fluorochrome emits green or orangish red fluorescence when it binds to bacterial DNA or RNA, respectively. We took advantage of the image by dividing the screen in several equal squares. For each screen containing an image divided in more than a 100 squares, we choose randomly a minimum of 30 squares to be counted.
Molecular identification
DNA was extracted from 1-mL cell cultures by the Wizard Genomic DNA Purification kit (Promega, USA) in accordance with the supplier’s instructions. Extraction products were visualized on 1% agarose gel with GelRed (Biotium, USA). Primers of 16S segment (forward 5′-CCTACGGGAGGCAGCAG-3′ and reverse 5′-GACTACCAGGGTATCTAATC-3′) were designed as previously described [2929 Ritchie LE, Steiner JM, Suchodolski JS: Assessment of microbial diversity along the feline intestinal tract using 16S rRNA gene analysis. FEMS Microbiol Ecol 2008, 66(3):590-598.]. DNA sample was amplified through polymerase chain reaction (PCR), which was performed in accordance with Ritchie et al.[2929 Ritchie LE, Steiner JM, Suchodolski JS: Assessment of microbial diversity along the feline intestinal tract using 16S rRNA gene analysis. FEMS Microbiol Ecol 2008, 66(3):590-598.], except for the primer annealing temperature, which was optimized for 58°C. PCR products (approximately 400-bp long) were analyzed on GelRed-stained 1% agarose gel, with Low DNA Mass Ladder (Invitrogen, Thermo Fisher Scientific, USA) as the molecular weight marker, and then purified by enzymes exonuclease I and shrimp alkaline phosphatase. Purified PCR products were sequenced in both directions by a MegaBACE 1000 automated sequencer (GE Healthcare, UK). The resulting chromatograms were analyzed and the DNA sequence was identified through the Basic Local Alignment Search Tool (BLAST–GenBank National Center for Biotechnology Information).
The 16S rRNA gene sequences used in the phylogenetic analysis were retrieved from the GenBank (accession numbers given in parentheses) as follows: P. aeruginosa YL84 (CP007147), P. alcaligenes LMG 1224 T (Z76653), P. citronellolis DSM 50332 T (Z76659), P. stutzeri ATCC 17589 (U25432), P. oleovorans DSM 1045 T (Z76665), P. flavescens B62 (U01916) and P. putida DSM 291 T (Z76667), and were aligned through the ClustalX program [3030 Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al.: Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23(21):2947-2948.]. An unrooted phylogenetic tree was constructed with the MEGA 6.0 program [3131 Tamura K, Stecher G, Peterson D, Filipski A, Kumar S: MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 2013, 30(12):2725-2729.] and the statistical Neighbor-Joining method[3232 Saitou N, Nei M: The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 1987, 4(4):406-425.] with proportional (p) distance was used.
All experimental research reported in this manuscript was in compliance with the Brazilian Ethics Committee for Animal tests.
Results
Samples of environmental water and sediment from the Patos Lagoon area including São Lourenço Beach were collected in order to isolate bacterial strains for the study of biodegradation of the hepatotoxin [D-Leu1] microcystin-LR. The analyses of water samples did not show meaningful differences except regarding the temperature; it was a slightly higher at spot P1 for several days.
Samples of water and sediment enabled the isolation of several strains (Table 1), whose ability for degrading microcystins were tested. However, most of them were not capable of degrading this cyanotoxin, except for the strain called DMXS.
The study of the biodegradation process of the hepatotoxin [D-Leu1] microcystin-LR was carried out in liquid medium. The decrease in the microcystin concentration in the medium is shown in Figure 4.
Decrease in the concentration of toxin [D-Leu1] microcystin-LR by the bacterial strain DMXS. ▲ Control, ■ Treatment.
For the controls, since the samples had been autoclaved and the toxin concentration did not decrease during the experiment, no physical, chemical or biological degradation was observed. Therefore, the bacterial community was inactivated in the autoclave process.
In the treatment flask containing the DMXS strain, 40% of the total toxin concentration occurred in three days. After 15 days, the toxin concentration was kept stable, around 60%. From the 15th day on, there was a steep decrease: from 60% to 0 in only four days (Figure 4).
The rate of toxin decrease was calculated and b was 0.054579 or 5 μg L−1/day. Half-life was 12.7 days. The equation used for the half-life rate was developed in accordance with Ozawa et al.[3333 Ozawa K, Yokoyama A, Ishikawa K, Kumagai M, Watanabe MF, Park HD: Accumulation and depuration of microcystin produced by the cyanobacterium Microcystis in a freshawater snail. Limnology 2003, 4(3):131-138.].
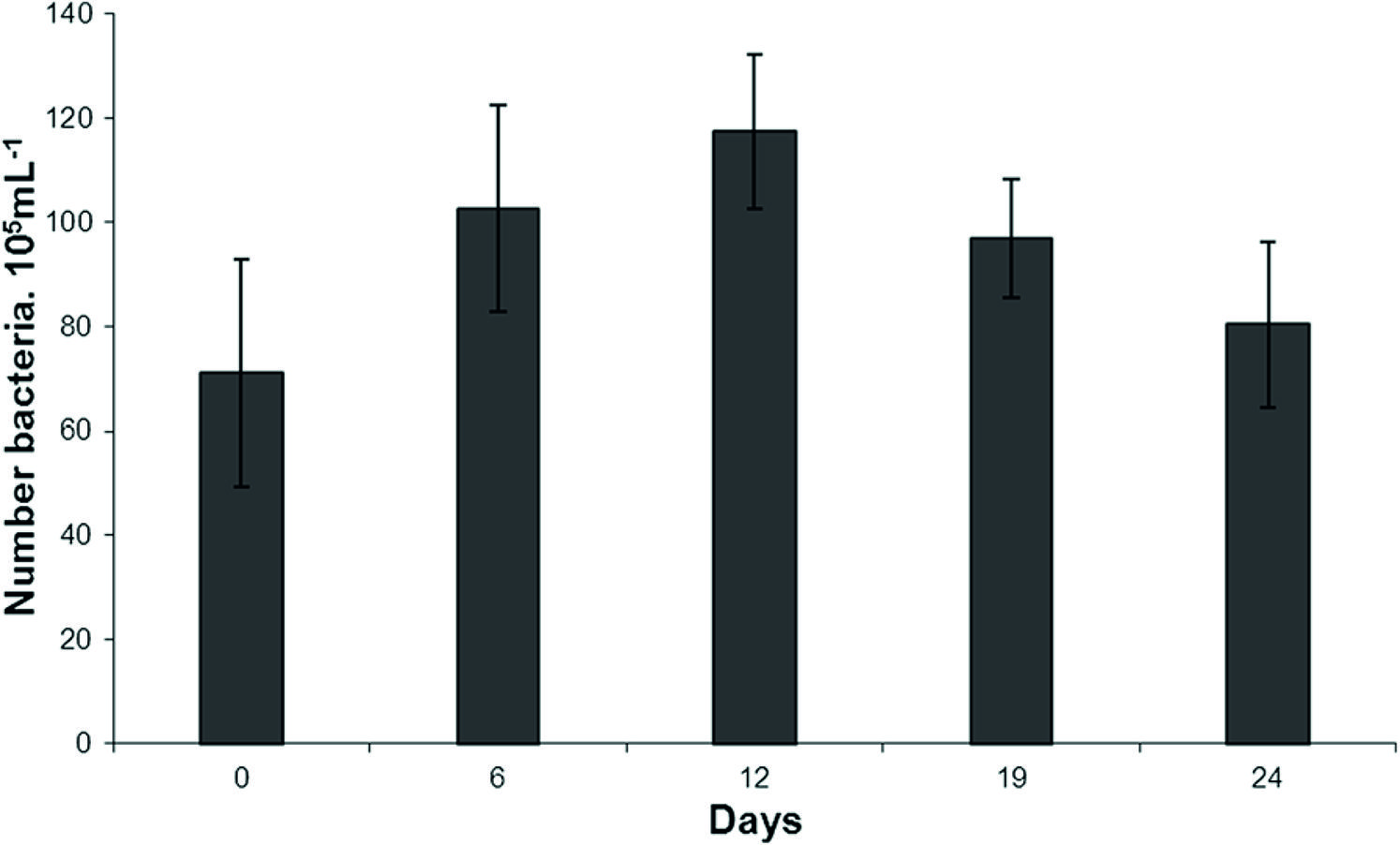
Figure 5 shows the growth of the strain DMXS in parallel with the uptake of the toxin during the same period of the experiment. The bacterial concentration increased up to the 12th day. The decrease started on the 19th day, when the toxin concentration started to decrease concomitantly.
In the inoculum, the bacterial concentration was 71 × 105 bacteria mL−1. It reached the highest concentration, 117 × 105 bacteria mL−1, on the 12th day.
In the presence of the toxin, the strain DMXS grew slowly during the experiment. The decrease in the growth started after it had reached its highest concentration, 117 × 105 bacteria.mL−1 when about 80% of the toxin had already been biotransformed.
The identification of the strain able of degrading the toxin was made by genetic sequencing; the total DNA was extracted from the isolated DMXS and the sequence of 16S rDNA was partially determined. The DNA sequence enabled the identification of the strain as Pseudomonas aeruginosaby the nucleotide BLAST tool (accession number HQ890467) and by the phylogenetic analysis using sequences from other species of the genus (Figure 6).
Phylogenetic tree of the isolated DMXS and other species of the genus Pseudomonas based on 16S rDNA sequence analysis.
Discussion
The waters in the hydrographic region of the Patos Lagoon have been studied and monitored regarding the presence and development of toxic cyanobacterial blooms as they are important for the economy and biodiversity in Rio Grande do Sul state, southern Brazil [1010 Yunes JS, Niencheski LF, Salomon P, Parise M, Beattie KA, Raggett SL, et al.: Effect of nutrient balance and physical factors on blooms of toxic cyanobacteria in the Patos Lagoon, southern Brazil. Verh Inter Verein Limnol 1998, 26:1796-1800.],[1212 Matthiensen A, Yunes JS, Codd GA: Ocorrência, distribuição e toxicidade de cianobactérias no estuário da Lagoa dos Patos. RS Rev Bras Biol 1999, 59(3):361-376.]. In an attempt to mitigate harmful effects of toxins on aquatic ecosystems, studies on the degradation and removal of cyanobacterial toxins have been carried out in several countries [1111 Lemes GA, Kersanach R, Pinto L da S, Dellagostin OA, Yunes JS, Matthiensen A:Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol Environ Saf 2008, 69(3):358-365.],[1616 Cousins IT, Bealing DJ, James HA, Sutton A: Biodegradation of microcystin-LR by indigenous mixed bacterial populations. Water Res 1996, 30(2):481-485.],[1717 Bourne DG, Jones GJ, Blakeley RL, Jones A, Negri AP, Riddles P: Enzymatic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin LR. Appl Environ Microbiol 1996, 62(11):4086-4094.],[3434 Jones GJ, Bourne DG, Blakeley RL, Doelle H: Degradation of the cyanobacterial hepatotoxin microcystin by aquatic bacteria. Nat Toxins 1994, 2(4):228-235.]-[3737 Edwards C, Graham D, Fowler N, Lawton LA: Biodegradation of microcystins and nodularin in freshwaters. Chemosphere 2008, 73(8):1315-1321.].
Biodegradation of the microcystin variant [D-Leu1] microcystin-LR was conducted in the laboratory using a strain isolated from sediment collected in the estuary. The strain DMXS was responsible for the decrease in the concentration of the toxin. It started right after the toxin was added to the experiment and was fast at the beginning. On the 19th day, toxin concentration was close to zero and, from the 24th day on, the toxin could not be identified anymore. The control was intact throughout the experiment; it means that the stability of the toxin was very high and that a possible contaminant bacterial community in the control was inactivated by the autoclave process.
The experiment was apparently stable for some time, from the 3rd to the 15th day, after the beginning of the toxin degradation process. However, degradation was slower only in this period. Other authors, including Chen et al.[1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.] and Edwards et al.[3737 Edwards C, Graham D, Fowler N, Lawton LA: Biodegradation of microcystins and nodularin in freshwaters. Chemosphere 2008, 73(8):1315-1321.], studied microcystin degradation and also found some similar results, i.g., some samples were only degraded after 10–20 days. The rate of toxin decrease was calculated and b was 0.054579 or 5 μg.L−1/day. Half-life was 12.7 days. Similar or higher values were also reported in other studies [1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.].
The organism responsible for the degradation of the toxin [D-Leu1] microcystin-LR is a bacterial strain isolated from sediment collected in the Patos Lagoon area (Rio Grande do Sul state, Brazil) at spot P1. Several studies of microcystin biodegradation have shown that there are bacteria in sediment collected in lakes [3838 Rapala J, Lahti K, Sivonen K, Niemelä SI: Biodegradability and adsorption on lake sediments of cyanobacterial hepatotoxins and anatoxin-a. Lett Appl Microbiol 1994, 19(6):423-428.]-[4040 Chen X, Yang X, Yang L, Xiao B, Wu X, Wang J, et al.: An effective pathway for the removal of microcystin LR via anoxic biodegradation in lake sediments. Water Res 2010, 44(6):1884-1892.]. This was observed both under aerobic [2121 Ishii H, Nishijima M, Abe T: Characterization of degradation process of cyanobacterial hepatotoxins by a gram-negative aerobic bacterium. Water Res 2004, 38(11):2667-2676.],[2222 Tsuji K, Asakawa M, Anzai Y, Sumino T, Harada KI: Degradation of microcystins using immobilized microorganism isolated in an eutrophic lake. Chemosphere 2006, 65(1):117-124.] or anoxic conditions [4040 Chen X, Yang X, Yang L, Xiao B, Wu X, Wang J, et al.: An effective pathway for the removal of microcystin LR via anoxic biodegradation in lake sediments. Water Res 2010, 44(6):1884-1892.].
During the experiment, the bacterial strain DMXS grew slowly even at high temperatures, but having the toxin [D-Leu1] microcystin-LR as its single carbon source. Temperatures between 27°C and 29°C are considered high in comparison with mean temperatures in the place where the strain was isolated. This strain metabolism is low and appropriate to this kind of environment in which temperatures do not vary much. Although there is abundance of bacteria in this place, the specific cell activities of sediment bacteria is low [4141 Haglund AL, Lantz P, Törnblom E, Tranvik L: Depth distribution of active bacteria and bacterial activity in lake sediment. FEMS Microbiol Ecol 2003, 46(1):31-38.].
It is also noteworthy that breaking the cyclic structure of microcystin molecules requires much energy from the bacterial metabolism. This may be a reason for the decrease on the rate of cell division and bacterial growth, as well.
The waters of Patos Lagoon harbor a vast diversity of bacteria [1313 Matthiensen A. Environmental and laboratory studies on the
properties and fate of microcystins from the cyanobacterium Microcystis. PhD
thesis. University of Dundee, UK; 2000:307. [available at University of Dundee
Library: http://www.dundee.ac.uk/library/search/].
http://www.dundee.ac.uk/library/search/...
]; however, as the present study has proven, the bacterial
population capable of degrading cyanotoxins is small. The present findings agree
with other studies that tested ability of bacteria from lake waters and sediment to
degrade microcystins in the laboratory. Some have reported that only 17% of the
strains are capable of carrying out the degradation [3535 Lahti K, Niemi RM, Rapala J, Sivonem K: Biodegradation of
cyanobacterial hepatotoxins–characterization of toxin degrading bacteria.
In Harmful Algae. Edited by Reguera B, Blanco J,
Fernández ML, Wyatt T. Xunta de Galicia and Intergovernmental Oceanographic
Commission of UNESCO, Santiago de Compostela; 1998.]. An Argentinean study evaluated three isolated strains, but only one
was capable of biodegrading microcystin-RR [4242 Amé MV, Echenique JR, Pflugmacher S, Wunderlin DA: Degradation
of Microcystin-RR by Sphingomonas sp. CBA4 isolated from San
Roque reservoir (Córdoba-Argentina). Biodegradation
2006, 17(5):447-455.].
Studies on microcystin biodegradation conducted in China with samples collected in Taihu Lake showed that only 17 out of 96 water samples were capable of carrying out the biodegradation. Twelve out of these 17 samples were collected near the sediment-water interface [1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.].
The capacity of an aquatic bacterium to degrade microcystins is not common, since only few species have the proper metabolism needed to do it [4343 Holst T, Jorgensen NOG, Jorgensen C, Johansen A: Degradation of microcystin in sediments at oxic and anoxic, denitrifying conditions. Water Res 2003, 37(19):4748-4760.]. The genus Sphingomonas has been pointed by most studies [1717 Bourne DG, Jones GJ, Blakeley RL, Jones A, Negri AP, Riddles P: Enzymatic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin LR. Appl Environ Microbiol 1996, 62(11):4086-4094.],[2121 Ishii H, Nishijima M, Abe T: Characterization of degradation process of cyanobacterial hepatotoxins by a gram-negative aerobic bacterium. Water Res 2004, 38(11):2667-2676.],[4444 Harada KI, Imanishi S, Kato H, Mizuno M, Ito E, Tsuji K: Isolation of Adda from microcystin-LR by microbial degradation. Toxicon 2004, 44(1):107-109.]-[4646 Saito T, Okano K, Park HD, Itayama T, Inamori Y, Neilan BA, et al.: Detection and sequencing of the microcystin LR-degrading gene, mlrA, from new bacteria isolated from Japanese lakes. FEMS Microbiol Lett 2003, 229(2):271-276.], even though other genera are also capable of degrading these toxins [1111 Lemes GA, Kersanach R, Pinto L da S, Dellagostin OA, Yunes JS, Matthiensen A:Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol Environ Saf 2008, 69(3):358-365.],[4747 Hu LB, Yang JD, Zhou W, Yin YF, Chen J, Shi ZQ: Isolation of a Methylobacillus sp. that degrades microcystin toxins associated with cyanobacteria. N Biotechnol 2009, 26(3–4):205-211.],[4848 Eleuterio L, Batista JR: Biodegradation studies and sequencing of microcystin-LR degrading bacteria isolated from a drinking water biofilter and a fresh water lake. Toxicon 2010, 55(8):1434-1442.].
Bacteria from sediment play an important role in the decomposition of organic matter and in geochemical cycles. These microorganisms are adapted to explore different sources of energy efficiently [66 Giovannardi S, Pollegioni L, Pomati F, Rossetti C, Sacchi S, Sessa L, et al.: Toxic cyanobacterial blooms in Lake Varese (Italy): a multidisciplinary approach. Environ Toxicol 1999, 14(1):127-134.],[4949 Nealson KH: Sediment bacteria: who’s there, what are they doing, and what’s new? Annu Rev Earth Planet Sci 1997, 25:403-434.]. Hence, microbial degradation has been recommended as an effective process to eliminate microcystins in fresh water [1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.]. In this sense, the water column and the sediment play an important role in the degradation of microcystins in aquatic environments [1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.],[3535 Lahti K, Niemi RM, Rapala J, Sivonem K: Biodegradation of cyanobacterial hepatotoxins–characterization of toxin degrading bacteria. In Harmful Algae. Edited by Reguera B, Blanco J, Fernández ML, Wyatt T. Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO, Santiago de Compostela; 1998.],[3737 Edwards C, Graham D, Fowler N, Lawton LA: Biodegradation of microcystins and nodularin in freshwaters. Chemosphere 2008, 73(8):1315-1321.]. Therefore low levels of microcystins in an environment may be due to bacterial degradation [1818 Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.].
Nowadays, biotechnological research on toxin biodegradation by sediment bacteria through removal of microcystins from water involves different techniques including: microorganisms immobilized in polyester resins, sand filters and bioreactors [2222 Tsuji K, Asakawa M, Anzai Y, Sumino T, Harada KI: Degradation of microcystins using immobilized microorganism isolated in an eutrophic lake. Chemosphere 2006, 65(1):117-124.],[3636 Bourne DG, Blakeley RL, Riddles P, Jones GJ: Biodegradation of the cyanobacterial toxin microcystin-LR in natural water and biologically active slow sand filters. Water Res 2006, 40(6):1294-1302.],[5050 Wang H, Ho L, Lewis DM, Brookes JD, Newcombe G: Discriminating and assessing adsorption and biodegradation removal mechanisms during granular activated carbon filtration of microcystins toxins. Water Res 2007, 41(18):4262-4270.]. These biotechnological applications–employing different aquatic microorganisms that biodegrade microcystins–are considered an efficient process to remove such contaminants from water [5050 Wang H, Ho L, Lewis DM, Brookes JD, Newcombe G: Discriminating and assessing adsorption and biodegradation removal mechanisms during granular activated carbon filtration of microcystins toxins. Water Res 2007, 41(18):4262-4270.]-[5252 Ho L, Hoefel D, Saint CP, Newcombe G: Isolation and identification of a novel microcystin-degrading bacterium from a biological sand filter. Water Res 2007, 41(20):4685-4695.].
Conclusion
The use of bacteria isolated from sediment for biotechnological applications is viable. Research has shown that sediment plays an important role as a source of bacteria capable of degrading cyanobacterial toxins. Results reported in this work corroborate previous studies and show that a bacterial strain from the genus Pseudomonas aeruginosa isolated from sediment collected in Patos Lagoon, in Brazil, biodegraded [D-Leu1] microcystin-LR in the laboratory.
Acknowledgments
The authors would like to thank CNPq, CAPES, FAURG for the funding of this research and their personal grants.
References
-
1Kotak BG, Kenefick SL, Fritz DL, Rousseaux CG, Prepas EE, Hrudey SE: Occurrence and toxicological evaluation of cyanobacterial toxins in Alberta lakes and farm dugouts. Water Res 1993, 27(3):495-506.
-
2Lawton LA, Codd GA: Cyanobacterial (Blue-Green Algal) toxins and their significance in UK and European waters. Water Environ J 1991, 5(4):460-465.
-
3Proença LAO, Tamanaha MS, Fonseca RS: Screening the toxicity and toxin content of blooms of the cyanobacterium Trichodesmium erythraeum (Ehrenberg) in northeast Brazil. J Venom Anim Toxins incl Trop Dis 2009, 15(2):204-215. [ http://www.scielo.br/scielo.php?pid=S1678-91992009000200004&script=sci_arttext ]
» http://www.scielo.br/scielo.php?pid=S1678-91992009000200004&script=sci_arttext -
4Yunes JS, Salomon PS, Matthiensen A, Beattie KA, Raggett SL, Codd GA: Toxic blooms of cyanobacteria in the Patos Lagoon Estuary, southern Brazil. J Aquat Ecosystem Health 1996, 5(4):223-229.
-
5Chorus I, Bartram J: Toxic cyanobacteria in water: A guide to their public health consequences, monitoring and management World Health Organization and E&FN Spon, London; 1999.
-
6Giovannardi S, Pollegioni L, Pomati F, Rossetti C, Sacchi S, Sessa L, et al.: Toxic cyanobacterial blooms in Lake Varese (Italy): a multidisciplinary approach. Environ Toxicol 1999, 14(1):127-134.
-
7Matthiensen A, Beattie KA, Yunes JS, Kaya K, Codd GA: [D-Leu1] Microcystin-LR, from the cyanobacterium Microcystis RST 9501 and from a Microcystis bloom in the Patos Lagoon estuary, Brazil. Phytochemistry 2000, 55(5):383-387.
-
8Jochimsen EM, Carmichael WW, An J, Cardo DM, Cookson ST, Holmes CEM, et al.: Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N Engl J Med 1998, 338:873-878.
-
9Pouria S, De Andrade A, Barbosa J, Cavalcanti RL, Barreto VT, Ward CJ, et al.: Fatal microcystin intoxication in haemodialysis unit in Caruaru. Brazil Lancet 1998, 352(9121):21-26.
-
10Yunes JS, Niencheski LF, Salomon P, Parise M, Beattie KA, Raggett SL, et al.: Effect of nutrient balance and physical factors on blooms of toxic cyanobacteria in the Patos Lagoon, southern Brazil. Verh Inter Verein Limnol 1998, 26:1796-1800.
-
11Lemes GA, Kersanach R, Pinto L da S, Dellagostin OA, Yunes JS, Matthiensen A:Biodegradation of microcystins by aquatic Burkholderia sp. from a South Brazilian coastal lagoon. Ecotoxicol Environ Saf 2008, 69(3):358-365.
-
12Matthiensen A, Yunes JS, Codd GA: Ocorrência, distribuição e toxicidade de cianobactérias no estuário da Lagoa dos Patos. RS Rev Bras Biol 1999, 59(3):361-376.
-
13Matthiensen A. Environmental and laboratory studies on the properties and fate of microcystins from the cyanobacterium Microcystis. PhD thesis. University of Dundee, UK; 2000:307. [available at University of Dundee Library: http://www.dundee.ac.uk/library/search/].
» http://www.dundee.ac.uk/library/search/ -
14Rinehart KL, Namikoshi M, Choi BW: Structure and biosynthesis of toxins from blue-green algae (cyanobacteria). J Appl Phycol 1994, 6(2):159-176.
-
15Sivonen K, Jones G: Cyanobacterial toxins. In Toxic cyanobacteria in water: A guide to their public health consequences, monitoring and management World Health Organization, E&FN Spon, London; 1999:41-111.
-
16Cousins IT, Bealing DJ, James HA, Sutton A: Biodegradation of microcystin-LR by indigenous mixed bacterial populations. Water Res 1996, 30(2):481-485.
-
17Bourne DG, Jones GJ, Blakeley RL, Jones A, Negri AP, Riddles P: Enzymatic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin LR. Appl Environ Microbiol 1996, 62(11):4086-4094.
-
18Chen W, Song L, Peng L, Wan N, Zhang X, Gan N: Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res 2008, 42(3):763-773.
-
19Ji RP, Lu XW, Li XN, Pu YP: Biological degradation of algae and microcystins by microbial enrichment on artificial media. Ecol Eng 2009, 35(11):1584-1588.
-
20Matthiensen A, Metcalf JS, Ferreira AHF, Yunes JS, Codd GA: Biodegradation and biotransformation of microcystin by aquatic microbes in estuarine waters from the Patos Lagoon, RS, Brazil. In Mycotoxins and Phycotoxins in perpective at the turn of the Millenium Edited by Koe WJ, Samson SA, Egmond HP, Gilbert J, Wageningen SM. Posen & Looyen, Brazil; 2000:527-36.
-
21Ishii H, Nishijima M, Abe T: Characterization of degradation process of cyanobacterial hepatotoxins by a gram-negative aerobic bacterium. Water Res 2004, 38(11):2667-2676.
-
22Tsuji K, Asakawa M, Anzai Y, Sumino T, Harada KI: Degradation of microcystins using immobilized microorganism isolated in an eutrophic lake. Chemosphere 2006, 65(1):117-124.
-
23Stanier RY, Kunisawa R, Mandel M, Cohen-Bazire G: Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol Rev 1971, 35(2):171-205.
-
24Lawton LA, Edwards C, Codd GA: Extraction and High-performance liquid chromatographic method for the determination of microcystins in raw and treated waters. Analyst (Cambridge, U K) 1994, 119:1525-1530.
-
25Morsen A, Rehm HJ: Degradation of phenol by mixed culture of Pseudomonas putida and Crytococcus elinonii adsorbed on activated carbon. Appl Microbiol Biotechnol 1987, 26:283-288.
-
26Hobbie JE, Daley RJ, Jasper S: Use of nuclepore filters for counting bacteria by fluorescence microscopy. Appl Environ Microbiol 1977, 33(5):1225-1228.
-
27Pomroy AJ: Direct counting of bacteria preserved with Lugol Iodine solution. Appl Environ Microbiol 1984, 47(5):1191-1192.
-
28Nishino SF: Direct acridine orange counting of bacteria preserved with acidified Lugol Iodine. Appl Environ Microbiol 1986, 52(3):602-604.
-
29Ritchie LE, Steiner JM, Suchodolski JS: Assessment of microbial diversity along the feline intestinal tract using 16S rRNA gene analysis. FEMS Microbiol Ecol 2008, 66(3):590-598.
-
30Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al.: Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23(21):2947-2948.
-
31Tamura K, Stecher G, Peterson D, Filipski A, Kumar S: MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 2013, 30(12):2725-2729.
-
32Saitou N, Nei M: The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 1987, 4(4):406-425.
-
33Ozawa K, Yokoyama A, Ishikawa K, Kumagai M, Watanabe MF, Park HD: Accumulation and depuration of microcystin produced by the cyanobacterium Microcystis in a freshawater snail. Limnology 2003, 4(3):131-138.
-
34Jones GJ, Bourne DG, Blakeley RL, Doelle H: Degradation of the cyanobacterial hepatotoxin microcystin by aquatic bacteria. Nat Toxins 1994, 2(4):228-235.
-
35Lahti K, Niemi RM, Rapala J, Sivonem K: Biodegradation of cyanobacterial hepatotoxins–characterization of toxin degrading bacteria. In Harmful Algae Edited by Reguera B, Blanco J, Fernández ML, Wyatt T. Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO, Santiago de Compostela; 1998.
-
36Bourne DG, Blakeley RL, Riddles P, Jones GJ: Biodegradation of the cyanobacterial toxin microcystin-LR in natural water and biologically active slow sand filters. Water Res 2006, 40(6):1294-1302.
-
37Edwards C, Graham D, Fowler N, Lawton LA: Biodegradation of microcystins and nodularin in freshwaters. Chemosphere 2008, 73(8):1315-1321.
-
38Rapala J, Lahti K, Sivonen K, Niemelä SI: Biodegradability and adsorption on lake sediments of cyanobacterial hepatotoxins and anatoxin-a. Lett Appl Microbiol 1994, 19(6):423-428.
-
39Rapala J, Berg KA, Lyra C, Niemi RM, Manz W, Suomalainen S, et al.: Paucibacter toxinivorans gen. nov., sp. nov., a bacterium that degrades cyclic cyanobacterial hepatotoxins microcystins and nodularin. Int J Syst Evol Microbiol 2005, 55(Pt 4):1563-1568.
-
40Chen X, Yang X, Yang L, Xiao B, Wu X, Wang J, et al.: An effective pathway for the removal of microcystin LR via anoxic biodegradation in lake sediments. Water Res 2010, 44(6):1884-1892.
-
41Haglund AL, Lantz P, Törnblom E, Tranvik L: Depth distribution of active bacteria and bacterial activity in lake sediment. FEMS Microbiol Ecol 2003, 46(1):31-38.
-
42Amé MV, Echenique JR, Pflugmacher S, Wunderlin DA: Degradation of Microcystin-RR by Sphingomonas sp. CBA4 isolated from San Roque reservoir (Córdoba-Argentina). Biodegradation 2006, 17(5):447-455.
-
43Holst T, Jorgensen NOG, Jorgensen C, Johansen A: Degradation of microcystin in sediments at oxic and anoxic, denitrifying conditions. Water Res 2003, 37(19):4748-4760.
-
44Harada KI, Imanishi S, Kato H, Mizuno M, Ito E, Tsuji K: Isolation of Adda from microcystin-LR by microbial degradation. Toxicon 2004, 44(1):107-109.
-
45Park HD, Sasaki Y, Maruyama T, Yanagisawa E, Hiraishi A, Kato K: Degradation of the cyanobacterial hepatotoxin microcystin by a new bacterium isolated from a hypertrophic lake. Environ Toxicol 2001, 16(4):337-343.
-
46Saito T, Okano K, Park HD, Itayama T, Inamori Y, Neilan BA, et al.: Detection and sequencing of the microcystin LR-degrading gene, mlrA, from new bacteria isolated from Japanese lakes. FEMS Microbiol Lett 2003, 229(2):271-276.
-
47Hu LB, Yang JD, Zhou W, Yin YF, Chen J, Shi ZQ: Isolation of a Methylobacillus sp. that degrades microcystin toxins associated with cyanobacteria. N Biotechnol 2009, 26(3–4):205-211.
-
48Eleuterio L, Batista JR: Biodegradation studies and sequencing of microcystin-LR degrading bacteria isolated from a drinking water biofilter and a fresh water lake. Toxicon 2010, 55(8):1434-1442.
-
49Nealson KH: Sediment bacteria: who’s there, what are they doing, and what’s new? Annu Rev Earth Planet Sci 1997, 25:403-434.
-
50Wang H, Ho L, Lewis DM, Brookes JD, Newcombe G: Discriminating and assessing adsorption and biodegradation removal mechanisms during granular activated carbon filtration of microcystins toxins. Water Res 2007, 41(18):4262-4270.
-
51Ho L, Gaudieux AL, Fanok S, Newcombe G, Humpage AR: Bacterial degradation of microcystin toxins in drinking water eliminates their toxicity. Toxicon 2007, 50(3):438-441.
-
52Ho L, Hoefel D, Saint CP, Newcombe G: Isolation and identification of a novel microcystin-degrading bacterium from a biological sand filter. Water Res 2007, 41(20):4685-4695.
Publication Dates
-
Publication in this collection
2015
History
-
Received
14 Apr 2014 -
Accepted
26 Jan 2015 -
Accepted
24 Feb 2015