Resumos
O comportamento de 20 genótipos de soja foi avaliado em relação à preferência para oviposição, atratividade para adultos e colonização das plantas pela mosca branca Bemisia tabaci (Genn.) biótipo B, em teste de livre escolha, em casa de vegetação. O experimento foi conduzido com plantas de soja nas fases vegetativa e reprodutiva, infestadas artificialmente com esse inseto. As cultivares IAC 17, IAC 19 e a linhagem D 75-10169 apresentaram resistência do tipo não preferência para oviposição, baixos níveis de colonização e atratividade para adultos em relação a B. tabaci biótipo B. Também em casa de vegetação, foi avaliada a densidade de tricomas na superfície abaxial de folíolos dos 20 genótipos de soja, nos estratos superior, médio e inferior das plantas, nas fases de desenvolvimento em que as plantas foram avaliadas no teste de livre escolha. A linhagem D 75-10169 apresentou baixa densidade de tricomas, enquanto a cultivar IAC Holambra Stwart mostrou-se altamente pubescente. Foram verificadas correlações entre atratividade para adultos e preferência para oviposição e entre atratividade para adultos e colonização das plantas. Para confirmação dos resultados de preferência para oviposição, foi realizado teste sem chance de escolha, em casa de vegetação, no qual foi avaliado o comportamento de cinco genótipos (um altamente suscetível e os quatro menos preferidos para oviposição no teste com chance de escolha). A linhagem PI 227687, embora apresente resistência múltipla a insetos, foi altamente ovipositada por B. tabaci biótipo B, enquanto que 'IAC 19', 'IAC 17', 'Coodetec 201' e a linhagem D 75-10169 mantiveram a resistência do tipo não-preferência para oviposição.
Glicyne max; resistência de plantas; preferência para oviposição; mosca branca
Twenty soybean genotypes were evaluated in relation to silverleaf whitefly [Bemisia tabaci (Genn.) biotype B] resistance, in a free-choice test, under greenhouse conditions. The experiment was carried out with artificially infested soybean plants and evaluated during vegetative and reproductive stages. Line D 75-10169 and cultivars IAC 17 and IAC 19 presented nonpreference for oviposition, weak colonization and low adult attractiveness. Trichome density at abaxial trifoliolate surface was also measured in twenty soybean genotypes, at three plant positions (upper, middle and lower canopies), at the same development stages of the free-choice test. The line D 75-10169 presented the lowest trichome density while the cultivar 'IAC Holambra Stwart' showed the highest one. Significant correlations were detected between adult attractiveness and oviposition preference and between adult attractiveness and colonization level. To confirm the oviposition preference results, a no-choice test was carried out, also in greenhouse conditions, and the performance of five soybean genotypes (one highly susceptible and four with the smallest levels of oviposition) was checked. The line PI 227687, though showing multiple resistance to insects, was highly oviposited by silverleaf whitefly, while 'IAC 19', 'IAC 17', 'Coodetec 201' and line D 75-10169 showed oviposition nonpreference as the mecanism of resistance. The results strongly suggest that the resistance observed in those genotypes is stable.
Glicyne max; host plant resistance; oviposition preference; silverleaf whitefly
CROP PROTECTION
Resistência de Genótipos de Soja a Bemisia tabaci (Genn.) Biótipo B (Hemiptera: Aleyrodidae)
GIULIANA ETORE DO VALLE1,2 E ANDRÉ L. LOURENÇÃO1
1Instituto Agronômico de Campinas (IAC), C. postal 28, 13001-970, Campinas, SP
2Pós-Graduanda, Bolsista da Capes
Resistance of Soybean Genotypes to Bemisia tabaci (Genn.) Biotype B (Hemiptera: Aleyrodidae)
ABSTRACT Twenty soybean genotypes were evaluated in relation to silverleaf whitefly [Bemisia tabaci (Genn.) biotype B] resistance, in a free-choice test, under greenhouse conditions. The experiment was carried out with artificially infested soybean plants and evaluated during vegetative and reproductive stages. Line D 75-10169 and cultivars IAC 17 and IAC 19 presented nonpreference for oviposition, weak colonization and low adult attractiveness. Trichome density at abaxial trifoliolate surface was also measured in twenty soybean genotypes, at three plant positions (upper, middle and lower canopies), at the same development stages of the free-choice test. The line D 75-10169 presented the lowest trichome density while the cultivar 'IAC Holambra Stwart' showed the highest one. Significant correlations were detected between adult attractiveness and oviposition preference and between adult attractiveness and colonization level. To confirm the oviposition preference results, a no-choice test was carried out, also in greenhouse conditions, and the performance of five soybean genotypes (one highly susceptible and four with the smallest levels of oviposition) was checked. The line PI 227687, though showing multiple resistance to insects, was highly oviposited by silverleaf whitefly, while 'IAC 19', 'IAC 17', 'Coodetec 201' and line D 75-10169 showed oviposition nonpreference as the mecanism of resistance. The results strongly suggest that the resistance observed in those genotypes is stable.
KEY WORDS: Glicyne max, host plant resistance, oviposition preference, silverleaf whitefly.
RESUMO O comportamento de 20 genótipos de soja foi avaliado em relação à preferência para oviposição, atratividade para adultos e colonização das plantas pela mosca branca Bemisia tabaci (Genn.) biótipo B, em teste de livre escolha, em casa de vegetação. O experimento foi conduzido com plantas de soja nas fases vegetativa e reprodutiva, infestadas artificialmente com esse inseto. As cultivares IAC 17, IAC 19 e a linhagem D 75-10169 apresentaram resistência do tipo não preferência para oviposição, baixos níveis de colonização e atratividade para adultos em relação a B. tabaci biótipo B. Também em casa de vegetação, foi avaliada a densidade de tricomas na superfície abaxial de folíolos dos 20 genótipos de soja, nos estratos superior, médio e inferior das plantas, nas fases de desenvolvimento em que as plantas foram avaliadas no teste de livre escolha. A linhagem D 75-10169 apresentou baixa densidade de tricomas, enquanto a cultivar IAC Holambra Stwart mostrou-se altamente pubescente. Foram verificadas correlações entre atratividade para adultos e preferência para oviposição e entre atratividade para adultos e colonização das plantas. Para confirmação dos resultados de preferência para oviposição, foi realizado teste sem chance de escolha, em casa de vegetação, no qual foi avaliado o comportamento de cinco genótipos (um altamente suscetível e os quatro menos preferidos para oviposição no teste com chance de escolha). A linhagem PI 227687, embora apresente resistência múltipla a insetos, foi altamente ovipositada por B. tabaci biótipo B, enquanto que 'IAC 19', 'IAC 17', 'Coodetec 201' e a linhagem D 75-10169 mantiveram a resistência do tipo não-preferência para oviposição.
PALAVRAS-CHAVE: Glicyne max, resistência de plantas, preferência para oviposição, mosca branca.
Moscas brancas (Hemiptera: Aleyrodidae) são insetos que sugam a seiva do floema das plantas hospedeiras, tanto na fase imatura como na adulta, podendo causar danos diretos e indiretos, além de transmitir viroses. Atualmente, são consideradas um grupo importantíssimo em âmbito mundial, veiculando mais de 40 fitoviroses diferentes, sendo as únicas transmissoras de geminivírus (Brown & Bird 1992). Uma das espécies mais importantes é Bemisia tabaci (Gennadius), hoje considerada um complexo (Brown et al. 1995), com amplo espectro de hospedeiras, criando-se em mais de 500 espécies de plantas, pertencentes a 74 famílias botânicas (Greathead 1986). Dependendo da cultura, época e nível de infestação, entre outros fatores, os prejuízos causados por essa mosca branca podem variar entre 20 e 100% (Brown & Bird 1992).
A existência de biótipos de B. tabaci foi proposta na década de 50, após a descoberta de populações morfologicamente semelhantes, que exibiam traços biológicos diferentes com relação à afinidade para a planta hospedeira, aos graus de sintomas de viroses, a resistência a inseticidas, a morfologia e ao comportamento dos insetos (Costa & Brown 1991, Bedford et al. 1994, Brown et al. 1995), tendo sido confirmada por estudos com marcadores de aloenzimas (Perring et al. 1992) e com RAPD-PCR (randomly amplified polymorphic DNA) (Gawel & Bartlett 1993).
Várias diferenças biológicas têm sido atribuídas aos dois biótipos de Bemisia: o biótipo B coloca significativamente mais ovos (Costa & Brown 1991), ingere maior quantidade de seiva da planta e excreta maior volume de honeydew (Byrne & Miller 1990), além de possuir maior gama de hospedeiros quando comparado ao biótipo A (Bedford et al. 1994).
No Brasil, B. tabaci biótipo B foi introduzida no início da década de 90. A partir de 1991, esta espécie de mosca branca passou a ser importante devido a altas infestações em plantas de interesse econômico. Lourenção & Nagai (1994) verificaram altas populações do biótipo B no Estado de São Paulo, na região de Campinas. Infestações severas foram observadas em plantas ornamentais e também em culturas de brócolis, berinjela, abóbora e tomate, nestas últimas duas causando, respectivamente, o prateamento das folhas e o amadurecimento irregular dos frutos, anomalias fisiológicas características desse biótipo. No Distrito Federal, as primeiras observações dessa espécie de mosca branca foram verificadas em tomateiro, em 1993 (França et al.1996). Posteriormente, o inseto alcançou as principais regiões agrícolas do país (Villas Bôas et al. 1997).
O controle de moscas brancas tem sido feito quase que exclusivamente por inseticidas e por tratos culturais. Prabhaker et al. (1985) mencionaram, entretanto, que características biológicas e comportamentais de B. tabaci como rápido desenvolvimento, alta fecundidade e grande capacidade de dispersão são fatores que aumentam a probabilidade de aparecimento de resistência aos inseticidas comerciais de diferentes grupos químicos (Prabhaker et al. 1989, Dittrich et al. 1990). Em razão desse e de outros problemas causados pelos inseticidas no agroecossistema, métodos alternativos de controle de moscas brancas vêm sendo estudados. Dentre esses métodos, pode-se citar a resistência de plantas a insetos. O estudo de resistência varietal a B. tabaci biótipo B é uma área que pode e deve ser explorada com o intuito de reduzir os danos causados por esse inseto (McAuslane 1996), apresentando grande potencial como estratégia de manejo em um programa integrado.
O presente trabalho foi realizado com o objetivo de avaliar genótipos de soja com diferentes graus de resistência a insetos com relação à infestação pela mosca branca B. tabaci biótipo B, a fim de determinar preferência para oviposição, atratividade para adultos e colonização das plantas nesse germoplasma. Também foi realizado um teste com o intuito de correlacionar a densidade dos tricomas presentes na superfície abaxial de folíolos com preferência para oviposição, atratividade para adultos e colonização das plantas.
Material e Métodos
Os experimentos foram conduzidos em casa de vegetação e em laboratório, no setor de Entomologia do Centro de Fitossanidade localizado no Núcleo Experimental de Campinas, no Instituto Agronômico de Campinas (IAC), durante o período de dezembro de 1999 a maio de 2001.
Criação Estoque de B. tabaci Biótipo B. A mosca branca B. tabaci biótipo B foi obtida em criação mantida no setor de Entomologia do IAC, sobre plantas de soja 'IAC 15-2', leiteiro (Euphorbia heterophylla L.) e poinsetia (Euphorbia pulcherrima Wild.). A referida espécie foi identificada pela Dra. Judith K. Brown, Universidade do Arizona, EUA.
Preferência Para Oviposição, Atratividade Para Adultos e Colonização das Plantas em Teste com Chance de Escolha. Foi estudada a preferência para oviposição de B. tabaci biótipo B em 20 genótipos de soja (Tabela 1), incluindo-se cultivares de expressão de plantio no Estado de São Paulo, cultivares IAC e as principais fontes de resistência a insetos utilizadas em programas de melhoramento, principalmente no IAC (Kogan 1989, Lourenção et al. 1989).
Para cultivo das plantas, foram utilizados vasos de alumínio (33 cm de diâmetro de boca x 31 cm de altura x 24 cm de diâmetro de base) com capacidade para 16 litros, preenchidos com mistura de terra e composto orgânico e mantidos em casa de vegetação. Sob os vasos, colocou-se um prato, periodicamente preenchido com água, de maneira a suprir as necessidades hídricas das plantas. Foram semeados nove vasos de cada genótipo, mantendo-se após o desbaste, duas plantas por vaso. Vasos de alumínio, com duas plantas de soja 'IAC 15-2' (15 cm de diâmetro de boca x 16 cm de altura x 10 cm de diâmetro de base), foram utilizados para infestação artificial. Após 24h de exposição dentro da criação estoque de B. tabaci biótipo B, os mesmos, com cerca de 2500 adultos de mosca branca cada, foram transferidos para a casa de vegetação, onde foi conduzido o experimento. Nessa ocasião, as plantas estavam no estágio vegetativo V2 (Fehr & Caviness 1977). Os vasos que serviram como focos de infestação foram dispostos de forma eqüidistante dos vasos com os genótipos, na proporção de 1:4, respectivamente, durante o período de uma semana.
A primeira avaliação foi realizada sete dias após a exposição aos adultos, quando as plantas encontravam-se nos estágios vegetativos V2 e V3. Foram avaliados dois folíolos por planta, tomando-se a terceira folha apical completamente desenvolvida, de cima para baixo, num total de quatro folíolos por parcela, nos quais efetuou-se a contagem do número de adultos presentes nas superfícies abaxiais. Em seguida, esses folíolos foram removidos e acondicionados em folhas de papel envoltas em saco plástico, levados para laboratório e armazenados em geladeira, para posterior manuseio. No laboratório, com auxílio de microscópio estereoscópico, foi contado o número de ovos na face abaxial de cada folíolo. Os folíolos removidos tiveram seu contorno desenhado em folhas de papel, recortados e medidos através de medidor de área foliar LI-COR (LI-3100A), para obtenção do número de ovos por 10 cm2 e de adultos por 10 cm2. Decorridos 15 dias da primeira avaliação, quando a maioria das plantas estava no estágio reprodutivo R1, foi feita nova avaliação, seguindo-se os procedimentos adotados para o estágio vegetativo. Duas semanas após, quando a maioria das plantas encontrava-se entre os estágios R1 a R4, foi feita a terceira avaliação, de forma semelhante às anteriores.
Um mês após o término da última avaliação, quando a maioria das plantas encontrava-se entre os estágios reprodutivos R4 e R7, estimou-se a colonização das plantas pela mosca branca nos diferentes genótipos estudados. Para isso, foi utilizada uma escala de notas, variando de 0 a 6, sendo: 0 = plantas com folíolos sem infestação; 1 = plantas com folíolos com poucos ovos e ninfas; ...; 6 = plantas com folíolos totalmente colonizados por ovos e ninfas (Fig. 1).
O delineamento empregado foi o de blocos casualizados, composto por 20 tratamentos e nove repetições, totalizando 180 parcelas. Cada parcela foi constituída por um vaso com duas plantas de soja e cada observação pela média de quatro folíolos, sendo dois de cada planta.
Densidade de Tricomas. Para esta avaliação, foram usados folíolos dos estratos superior, médio e inferior das plantas, nos estágios de desenvolvimento nos quais foram feitas as avaliações no teste de preferência para oviposição e de atratividade para adultos, com livre chance de escolha. A densidade foi medida pela contagem dos tricomas presentes numa área de 16 mm2, na face abaxial de folíolos dos 20 genótipos de soja, sendo os folíolos examinados com auxílio de microscópio estereoscópico, sob aumento de 32X. Utilizaram-se oito folíolos de cada terço da planta (estratos inferior, médio e superior), totalizando 24 folíolos para cada genótipo. O delineamento foi inteiramente casualizado, com 20 tratamentos e oito repetições, totalizando 160 parcelas, para cada estrato da planta. Cada parcela foi constituída por um folíolo de soja e cada observação pela média do número de tricomas presentes nas porções basal e apical de cada folíolo, procedimento similar ao adotado por McAuslane (1996).
Preferência Para Oviposição em Teste sem Chance de Escolha. Foi avaliada nos quatro genótipos que apresentaram as menores médias de número de ovos/10 cm2 e no genótipo mais suscetível, observados no teste com chance de escolha: 'IAC 17', 'IAC 19', D 75-10169, PI 227687 e 'Coodetec 201'. Utilizou-se o mesmo procedimento do teste anterior, adotando-se também o mesmo tipo de vaso. Para confinamento das moscas brancas, foram utilizadas gaiolas com armação de ferro de 35 cm de diâmetro x 70 cm de altura, cobertas com tecido voil. Quando as plantas estavam no estágio vegetativo V2 e V3, foi feita infestação artificial, colocando-se cerca de 200 adultos de B. tabaci biótipo B por vaso, durante quatro dias, conforme descrito por Oriani & Lara (2000). Quatro dias após a infestação, quando a maioria das plantas encontrava-se no estágio V3, avaliou-se a preferência para oviposição, considerando-se os procedimentos adotados no teste com chance de escolha, que incluíram contagem de ovos e medição de área foliar.
O delineamento utilizado foi blocos casualizados, composto por cinco tratamentos e dez repetições, totalizando 50 parcelas. Cada parcela foi constituída de um vaso com duas plantas de soja e cada observação foi obtida pela média de quatro folíolos, sendo dois de cada planta.
Análise Estatística. Quanto ao teste com chance de escolha, realizou-se a análise da variância, sendo as médias de número de ovos/10 cm2, colonização das plantas e número de tricomas/16 mm2 transformadas em  e as de número de adultos/10 cm2 em
e as de número de adultos/10 cm2 em  . No teste sem chance de escolha, as médias de número de ovos/10 cm2 foram transformadas em . Essas médias foram comparadas pelo teste de Tukey (P£0,05), utilizando o pacote estatístico SAS.
. No teste sem chance de escolha, as médias de número de ovos/10 cm2 foram transformadas em . Essas médias foram comparadas pelo teste de Tukey (P£0,05), utilizando o pacote estatístico SAS.
A correlação foi determinada separadamente para cada época de avaliação, sendo analisadas as seguintes variáveis: número de ovos/10 cm2, número de adultos/10 cm2 e média de tricomas totais na planta, sendo utilizado o pacote estatístico SAS. Utilizando a média das três avaliações, também foi determinada, conjuntamente, a correlação entre as variáveis: média de número de ovos/10 cm2, média de número de adultos/10 cm2, média do número de tricomas totais na planta e notas de colonização das plantas.
Resultados e Discussão
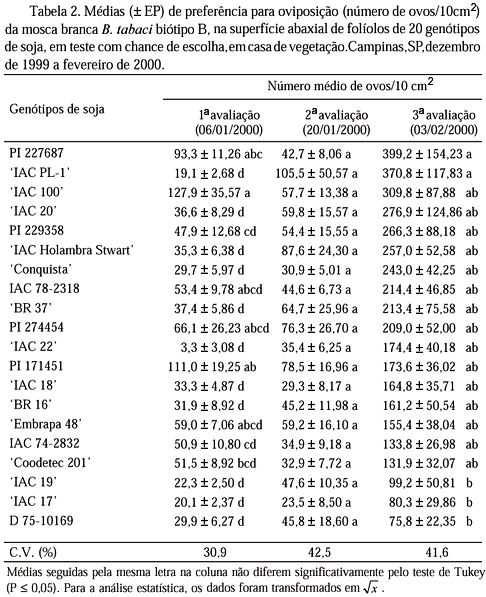
Preferência Para Oviposição em Teste com Chance de Escolha. Na primeira avaliação (Tabela 2), onze genótipos ('IAC PL-1', 'IAC 17', 'IAC 19', 'Conquista', D 75-10169, 'BR 16', 'IAC 22', 'IAC 18', 'IAC Holambra Stwart', 'IAC 20' e 'BR 37') apresentaram as menores médias de oviposição, diferindo significativamente de 'IAC 100', PI 171451 e PI 227687, que foram os mais ovipositados. Na segunda avaliação, as médias de oviposição variaram entre 23,5 ovos/10 cm2 ('IAC 17') e 105,5 ovos/10 cm2 ('IAC PL-1'); entretanto, não foram detectadas diferenças entre os genótipos. Na terceira avaliação, ocorreram as diferenças mais acentuadas entre os genótipos, provavelmente devido às altas infestações aos quais esses genótipos foram submetidos, pois nesta época já havia ocorrido a reinfestação natural dos insetos dentro da casa de vegetação. Verifica-se a menor oviposição em 'IAC 19', 'IAC 17' e D 75-10169 em comparação a PI 227687 e 'IAC PL-1'. Os resultados da PI 227687 ratificam aqueles obtidos por Rossetto et al. (1977), que também observaram alta preferência de B. tabaci para oviposição nesse genótipo.
Atratividade Para Adultos. Na primeira avaliação (Tabela 3), verifica-se que 'IAC PL-l', 'IAC 17', 'IAC 19', D 75-10169, 'Conquista'e IAC 78-2318 foram menos atrativos que PI 171451, 'IAC 100'e PI 227687, os quais apresentaram as maiores médias de adultos, ficando os demais genótipos em posição intermediária. Na segunda avaliação, observa-se que 'IAC 19', 'IAC 18' e 'IAC 17' foram os menos atrativos, diferindo de PI 227687, PI 274454 e 'IAC 20', que apresentaram as maiores médias. Na última avaliação, 'IAC 19' foi o genótipo menos atrativo, diferindo significativamente de 'IAC Holambra Stwart', PI 227687, 'BR 16', 'BR 37', 'IAC PL-1'e 'IAC 22', que tiveram as maiores médias de adultos. Ainda, foi nesta avaliação que se constataram as maiores médias de adultos, decorrentes da reinfestação natural dos insetos dentro da casa de vegetação.
'IAC 19', 'IAC 17' e D 75-10169 (Tabela 3), além de se situarem entre os genótipos menos atrativos aos adultos, também foram os genótipos menos preferidos para oviposição por B. tabaci biótipo B. Por outro lado, PI 227687, que ficou entre os mais atrativos, também foi o mais preferido para oviposição (Tabela 2).
Colonização. Também para este critério (Tabela 4), verifica-se o comportamento de resistência de 'IAC 19', D 75-10169 e 'IAC 17', que apresentaram as menores notas de colonização, diferindo dos demais genótipos, com exceção de 'BR 37', PI 229358, 'Coodetec 201', IAC 78-2318 e 'IAC 18'.
Lourenção & Miranda (1987) verificaram que, embora tivesse número relativamente alto de ovos de B. tabaci em testes de infestação com livre chance de escolha, a linhagem IAC 78-2318 apresentava baixa colonização por esse inseto, o que é confirmado pelos presentes resultados.
A avaliação da colonização das plantas pela mosca branca B. tabaci biótipo B vem ressaltar a resistência de 'IAC 19', 'IAC 17' e da linhagem D 75-10169, que já haviam demonstrado comportamento de resistência quanto à preferência para oviposição e à atratividade de adultos.
Observa-se que o genótipo mais suscetível à colonização, a cultivar Embrapa-48, situou-se no teste de preferência para oviposição entre aqueles de comportamento intermediário, sugerindo que seja um genótipo bastante favorável ao desenvolvimento das ninfas.
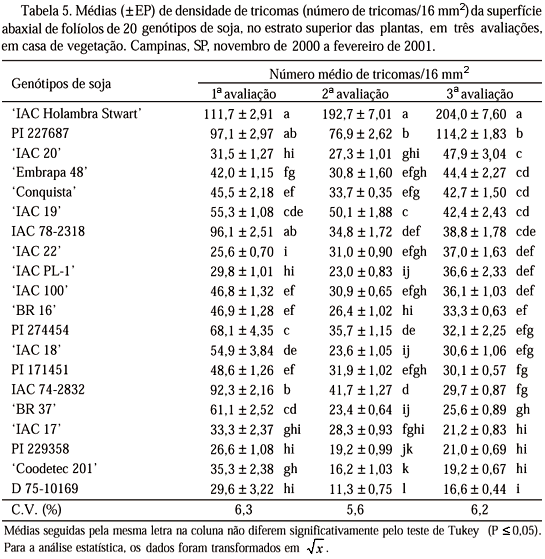
Densidade de Tricomas. Como as diferenças mais acentuadas observadas entre os genótipos quanto à preferência para oviposição ocorreram na terceira avaliação, a densidade de tricomas foi relacionada principalmente a essa época de avaliação. Com relação à densidade de tricomas no estrato superior das plantas (Tabela 5), observa-se que, na primeira avaliação, 'IAC 22', PI 229358, D 75-10169, 'IAC PL-1' e 'IAC 20' apresentaram as menores médias, diferindo dos demais, com exceção de 'IAC 17' e 'Coodetec 201'. Na segunda avaliação, a linhagem D 75-10169 apresentou a menor média, diferindo de todos os outros genótipos. 'IAC Holambra Stwart' e PI 227687 foram os genótipos com maiores médias de densidade de tricomas. Na terceira avaliação, D 75-10169, 'Coodetec 201', PI 229358 e 'IAC 17' apresentaram as menores médias, diferindo, com exceção de 'BR 37', de todos os demais. 'IAC Holambra Stwart' e PI 227687 apresentaram as maiores médias, o que já havia sido verificado nas avaliações anteriores para este estrato das plantas.
O elevado número de ovos/10 cm2 colocado por B. tabaci biótipo B na PI 227687, em teste com chance de escolha (Tabela 2), pode estar associado ao alto número de tricomas presentes nos folíolos desse genótipo (Tabela 5). McAuslane (1996) já havia verificado que genótipos de soja com alta densidade de tricomas são preferidos para oviposição por esse inseto em relação a genótipos glabros. De forma semelhante, pode-se associar a menor preferência para oviposição por B. tabaci biótipo B na linhagem D 75-10169 com a densidade de tricomas na superfície abaxial dos seus folíolos, uma vez que a linhagem apresentou as menores médias de tricomas.
Os tricomas são fortemente correlacionados com densidades de insetos e danos por alimentação para diferentes pragas. A alta densidade de tricomas, em algodoeiro (Berlinger 1986, Butter & Vir 1989), em soja (McAuslane et al. 1995, McAuslane et al. 1996) e em tomateiro (Heinz & Zalom 1995) está positivamente correlacionada com a oviposição de B. tabaci.
Butter & Vir (1989) sugerem que genótipos altamente pilosos podem fornecer um microclima mais adequado para a oviposição das fêmeas de B. tabaci. Além disso, fêmeas de B. tabaci preferem colocar os ovos na base de inserção dos tricomas (Omram & El-Khidir 1978, Berlinger 1986), fato que poderia estar associado a uma resposta evolutiva da pressão de seleção exercida por predadores e parasitóides (Butter & Vir 1989), já que estes são mais eficientes em folhas glabras ou com menor número de tricomas (Li et al. 1987). Ninfas fixadas na base de inserção dos tricomas, podem sofrer menor mortalidade decorrente da predação, além de conferir alta adaptação para fêmeas que exibam esse comportamento (Heinz & Zalom 1995).
Entretanto, 'IAC Holambra Stwart' foi muito atrativo aos adultos, apresentou a mais alta densidade de tricomas, mas situou-se de forma intermediária entre os genótipos com relação à preferência para oviposição. Em geral, variedades pubescentes são preferidas até certo nível, a partir do qual a pubescência começa a interferir na alimentação e na oviposição. Butler Jr. et al. (1991) observaram que houve declínio da população do inseto em algodoeiros com mais de 70 tricomas por 13,7 mm2. Isto poderia explicar o que ocorreu com essa cultivar, interferindo na oviposição.
Na verdade, o aumento da taxa de oviposição da mosca branca com o aumento da densidade de tricomas não é uma tendência constante para todas as plantas hospedeiras. A pubescência das folhas está negativamente correlacionada com a população de mosca branca em várias cucurbitáceas (McCreight & Kishaba 1991) e em Lagenaria siceraria (Molina) (Kishaba et al. 1992).
Análise de Correlação. Para a primeira avaliação (Tabela 6), os valores dos coeficientes demonstraram correlação altamente significativa entre número de ovos/10 cm2 e de adultos/10 cm2. Para a segunda avaliação, os valores dos coeficientes de correlação não foram significativos. Para a terceira avaliação, os valores dos coeficientes revelaram correlação significativa entre número de adultos/10 cm2 e média do número de tricomas totais na planta. Para a média das três avaliações, os valores dos coeficientes mostraram correlação significativa entre média do número de ovos/10 cm2 e de adultos/10 cm2 e entre média do número de adultos/10 cm2 e notas de colonização das plantas; portanto, verifica-se que existe associação positiva entre a atratividade para adultos e a preferência para oviposição e também entre atratividade para adultos e colonização das plantas.
Preferência Para Oviposição em Teste sem Chance de Escolha. Observa-se que PI 227687 foi o genótipo mais preferido para oviposição, diferindo significativamente dos demais, 'IAC 19', 'Coodetec 201', 'IAC 17' e D 75-10169, os quais não diferiram entre si (Tabela 7). Entre esses quatro genótipos, apenas 'IAC 19' apresentou densidade de tricomas próxima aos mais pubescentes (Tabela 5).
Embora o efeito dos tricomas na preferência para oviposição seja menos pronunciado em testes de confinamento (McAuslane 1996), 'IAC 19', 'Coodetec 201', 'IAC 17' e D 75-10169 já haviam sido os genótipos menos ovipositados no teste com chance de escolha, indicando que esses materiais mantém a característica de resistência, mesmo em condições em que o inseto não tem outro genótipo para ovipositar. Observações nesse sentido haviam sido feitas por Lourenção & Yuki (1982) para oviposição de B. tabaci em plantas de soja, em que os materiais mais resistentes mantinham essa característica em teste sem chance de escolha. Por outro lado, Elsey & Farnham (1994), trabalhando com B. oleraceae, e Blua et al. (1995), com abobrinha, beterraba e repolho, sugeriram que, em teste sem chance de escolha, as plantas perdem a característica de resistência, permitindo que as fêmeas de B. tabaci explorem novos hospedeiros.
As linhagens PI 171451, PI 227687 e PI 229358, que proporcionaram grande impulso nos estudos de resistência de soja a insetos após serem identificadas como portadoras de resistência múltipla (Van Duyn et al. 1971, Clark et al. 1972, Smith 1985, Beach & Todd 1988), apresentaram comportamento diferenciado frente a B. tabaci biótipo B. PI 227687 mostrou ser altamente preferida para oviposição, enquanto as outras duas introduções situaram-se entre os genótipos de comportamento intermediário. Rossetto et al. (1977), avaliando essas introduções em comparação com cultivares de interesse comercial naquela época frente a B. tabaci, também verificaram comportamento de suscetibilidade em PI 227687, enquanto PI 171451 e PI 229358 mostraram-se resistentes. Posteriormente, Lourenção & Yuki (1982) e Lourenção & Miranda (1987) confirmaram a menor oviposição de B. tabaci nessas duas introduções. Como no presente estudo PI 171451 e PI 229358 não exibiram comportamento de resistência frente à oviposição de mosca branca em níveis similares aos verificados por aqueles autores, pode-se atribuir esse fato ao germoplasma usado naquelas ocasiões, diferente do atual, e também ao biótipo de B. tabaci, B no presente caso, reconhecidamente mais nocivo à agricultura que aquele. Ainda, essas diferenças de comportamento entre as três PI's vêm ratificar relato de Kilen & Lambert (1986), que afirmaram que a resistência a insetos em cada uma dessas PI's é governada por diferentes fatores.
Com relação a outras fontes de resistência a insetos, verifica-se que 'IAC 100', cultivar resistente a percevejos (Rossetto et al. 1989), PI 274454, resistente a percevejos e a Omiodes indicata (Lourenção et al. 1985) e IAC 74-2832, apresentaram comportamento intermediário frente à oviposição e à colonização de B. tabaci biótipo B. Lourenção & Miranda (1987) verificaram que PI 274454 foi o genótipo mais ovipositado, além de apresentar alta colonização por B. tabaci, mostrando suscetibilidade a esse inseto.
Enfocando-se os genótipos que se destacaram pela resistência tanto a oviposição como pela atratividade para adultos e colonização por B. tabaci biótipo B, 'IAC 17' é uma cultivar que foi desenvolvida pelo IAC para resistência a insetos, sendo originada a partir do cruzamento envolvendo a linhagem D 72-9601, resistente a lagartas e derivada da PI 229358, o mesmo ocorrendo para 'IAC 19'. Essas duas cultivares apresentaram colonização por B. tabaci biótipo B visivelmente menos intensa que outras, como 'Iguaçu', em plantios comerciais de soja em Guaíra e Miguelópolis, durante o ano agrícola de 1997/98, época em que ocorreram surtos populacionais dessa mosca branca nessa região (A.L. Lourenção & M.A.C. Miranda, dados não publicados), o que vem corroborar os presentes resultados. A linhagem D 75-10169, da mesma forma que 'IAC 17' e 'IAC 19', tem como um dos parentais a PI 229358 e é resistente a insetos; com os resultados de resistência obtidos frente a B. tabaci biótipo B, pode ser considerada como uma boa fonte de resistência a insetos a ser utilizada em programas de melhoramento de soja.
Assim, é possível o uso imediato de 'IAC 17' e 'IAC 19' para regiões e/ou épocas em que B. tabaci biótipo B constitui problema na produção de soja ou mesmo em propriedades em que outras plantas afetadas direta ou indiretamente por essa mosca branca, como feijão, tomate, abóbora, hortaliças, ornamentais, são cultivadas simultaneamente ou em seqüência a essa leguminosa. O plantio dessas cultivares, aliado a outras técnicas de controle, como uso de inseticidas altamente seletivos, deverá diminuir de forma significativa as populações dessa mosca branca, beneficiando a soja contra danos diretos dessa praga, e outras culturas que são afetadas tanto por danos diretos como pela transmissão de vírus.
Literatura Citada
Received 06/07/01. Accepted 06/01/2002.
- Beach, R.M. & J.W. Todd. 1988. Foliage consumption and developmental parameters of the soybean looper and the velvetbean caterpillar (Lepidoptera: Noctuidae) reared on susceptible and resistant soybean genotypes. J. Econ. Entomol. 81: 310-316.
- Bedford, I.D., R.W. Briddon, J.K. Brown, R.C. Rosell & P.G. Markham. 1994. Geminivirus transmission and biological characterisation of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Appl. Biol. 125: 311-325.
- Berlinger, M.J. 1986. Host plant resistance to Bemisia tabaci Agric. Ecosyst. Environ. 17: 69-82.
- Blua, M.J., H.A. Yoshida & N.C. Toscano. 1995. Oviposition preference of two Bemisia species (Homoptera: Aleyrodidae). Environ. Entomol. 24: 88-93.
- Brown, J.K., D.R. Frolich & R.C. Rosell. 1995. The sweetpotato or silverleaf whiteflies: biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol. 40: 511-534.
- Brown, J.K. & J. Bird. 1992. Whitefly-transmitted geminiviruses and associated disorders in the Americas and Caribbean Basin. Plant Dis. 76: 220-225.
- Butler Jr, G.D., F.D. Wilson & G. Fishler. 1991. Cotton leaf trichomes and populations of Empoasca lybica and Bemisia tabaci Crop Prot. 10: 461-464.
- Butter, N.S. & B.K. Vir. 1989. Morphological basis of resistance in cotton to the whitefly Bemisia tabaci Phytoparasitica 17: 251-261.
- Byrne, D.N. & W.B. Miller. 1990. Carbohydrate and amino acid composition of phloem sap and honeydew produced by Bemisia tabaci J. Ins. Physiol. 36: 433-439.
- Clark, W.J., F.A. Harris, F.G. Maxwell & E.E. Hartwig. 1972. Resistance of certain soybean cultivars to bean leaf beetle, striped blister beetle, and bollworm. J. Econ. Entomol. 65: 1669-1672.
- Costa, H.S. & J.K. Brown. 1991. Variation in biological characteristics and esterase patterns among populations of Bemisia tabaci, and the association of one population with silverleaf symptom induction. Entomol. Exp. Appl. 61: 211-219.
- Dittrich, V., G.H. Ernst, O. Ruesch & S. UK. 1990. Resistance mechanisms in sweetpotato whitefly (Homoptera: Aleyrodidae) populations from Sudan, Turkey, Guatemala, and Nicaragua. J. Econ. Entomol. 83: 1665-1670.
- Elsey, K.D. & M.W. Farnham. 1994. Response of Brassica oleracea L. to Bemisia tabaci (Gennadius). HortScience 29: 814-817.
- Fehr, W.R. & C.E. Caviness. 1977. Stages of soybean development. Iowa Coop. Ext. Serv. Spec. Rep. 80. 12p.
- França, F.H., G.L. Villas Bôas & M.C. Branco. 1996. Ocorrência de Bemisia argentifolii Bellows & Perring (Homoptera: Aleyrodidae) no Distrito Federal. An. Soc. Entomol. Brasil 25: 369-372.
- Gawell, N.J. & A.C. Bartlett. 1993. Characterization of differences between whiteflies using RAPD-PCR. Ins. Mol. Biol. 2: 33-38.
- Greathead, A.H. 1986. Host plants, p.17-25. In M.J.W. Cock (ed.), Bemisia tabaci - a literature survey on the cotton whitefly with an annotated bibliography. Ascot, FAO/CAB, 121p.
- Heinz, K.M. & F.G. Zalom. 1995. Variation in trichome-based resistance to Bemisia argentifolii (Homoptera: Aleyrodidae) oviposition on tomato. J. Econ. Entomol. 88: 1494-1502.
- Kilen, T.C. & L. Lambert. 1986. Evidence for different genes controlling insect resistance in three soybean genotypes. Crop Sc. 26: 869-871.
- Kishaba, A.N., S. Castle, J.D. McCreight & P.R. Desjardins. 1992. Resistance of white-flowered gourd to sweetpotato whitefly. HortScience 27: 1217-1221.
- Kogan, M. 1989. Plant resistance in soybean insect control, p.1519-1525. In A.J. Pascale (ed.), World Soybean Research Conference IV. Buenos Aires: Orientácion Gráfica Editora, S.R.L., 1605p.
- Li, Z.H., F. Lammes, J.C. van Lenteren, P.W.T. Huisman, A. van Vianen & O.M.B. de Ponti. 1987. The parasite-host relationship between Encarsia formosa Gahan (Hymenoptera, Aphelinidae) and Trialeurodes vaporariorum (Westwood) (Homoptera: Aleyrodidae). XXV. Influence of leaf structure on the searching activity of Encarsia formosa J. Appl. Entomol. 104: 297-304.
- Lourenção, A.L., A.S. Costa & M.A.C. de Miranda. 1989. Sources of resistance to insect pests and virus vectors in the soybean germplasm tested at the Instituto Agronômico, Brazil, p.1578-1581. In: A.J. Pascale (ed.), World Soybean Research Conference IV. Buenos Aires: Orientácion Gráfica Editora S.R.L.1605p.
- Lourenção, A.L. & A.V. Yuki. 1982. Oviposição de Bemisia tabaci (Genn.) (Homoptera: Aleyrodidae) em três variedades de soja. Bragantia 41: 199-202.
- Lourenção, A.L., C.J. Rossetto & M.A.C. de Miranda. 1985. Resistência de soja a insetos. IV. Comportamento de cultivares e linhagens em relação a Hedylepta indicata (Fabr.). Bragantia 44: 149-157.
- Lourenção, A.L. & H. Nagai. 1994. Surtos populacionais de Bemisia tabaci no Estado de São Paulo. Bragantia 53: 53-59.
- Lourenção, A.L. & M.A.C. de Miranda. 1987. Resistência de soja a insetos: VIII. IAC 78-2318, linhagem com resistência múltipla. Bragantia 46: 65-72.
- McAuslane, H.J. 1996. Influence of leaf pubescence on ovipositional preference of Bemisia argentifolii (Homoptera: Aleyrodidae) on soybean. Environ. Entomol. 25: 834-841.
- McAuslane, H.J., F.A. Johnson, D.L. Colvin & B. Sojack. 1995. Influence of foliar pubescence on abundance and parasitism of Bemisia argentifolii (Homoptera: Aleyrodidae) on soybean and peanut. Environ. Entomol. 24: 1135-1143.
- McAuslane, H.J., S.E. Webb & G.W. Elmstrom. 1996. Resistance in germplasm of Cucurbita pepo to silverleaf, a disorder associated with Bemisia argentifolii (Homoptera: Aleyrodidae). Fla. Entomol. 79: 206-221.
- McCreight, J.D. & A.N. Kishaba. 1991. Reaction of cucurbit species to squash leaf curl virus and sweetpotato whitefly. J. Am. Soc. Hortic. Sci. 116: 137-141.
- Omram, H.H. & E. El-Khidir. 1978. On the preference of Bemisia tabaci Genn. (Homoptera: Aleyrodidae) on various cotton cultivars in Cukurova, Turkey. Agric., Ecosyst. Environ. 17: 83-88.
- Oriani, M.A. de G. & F.M. Lara. 2000. Oviposition preference of Bemisia tabaci (Genn.) Biotype B (Homoptera: Aleyrodidae) for Bean Genotypes Containing Arcelin in the Seeds. An. Soc. Entomol. Brasil 29: 565-572.
- Perring, T.M., A. Cooper, A. & D.J. Kazmer. 1992. Identification of the poinsettia strain of Bemisia tabaci (Homoptera: Aleyrodidae) on broccoli by electrophoresis. J. Econ. Entomol. 85: 1278-1284.
- Prabhaker, N., D.L. Coudriet & D.E. Meyerdirk. 1985. Insecticide resistance in the sweetpotato whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 78: 748-752.
- Prabhaker, N., N.C. Toscano & D.L. Coudriet. 1989. Susceptibility of the immature and adult stages of the sweet potato whitefly (Homoptera: Aleyrodidae) to selected insecticides. J. Econ. Entomol. 82: 983-988.
- Rossetto, C.J., O. Tisseli Fo, J. Cione, P.B. Gallo, L.F. Razera, J.P.F. Teixeira & N. Bortoletto. 1989. Soja 'IAC-100'. Agronômico 41: 2.
- Rossetto, D., A.S. Costa, M.A.C. de Miranda, V. Nagai & E. Abramides. 1977. Diferenças na oviposição de Bemisia tabaci em variedades de soja. An. Soc. Entomol. Brasil 6: 256-263.
- Smith, C.M. 1985. Expressions, mechanisms and chemistry of resistance in soybean, Glycine max L. (Merr.) to the soybean looper, Pseudoplusia includens (Walker). Insect Sc. Appl. 6: 243-248.
- Van Duyn, J.W., S.G. Turnipseed & J.D. Maxwell. 1971. Resistance in soybeans to the Mexican bean beetle. I. Sources of resistance. Crop Sci. 11: 572-573.
- Villas Bôas, G.L., F.H. França, A.C. de Ávila & I.C. Bezerra. 1997. Manejo integrado da mosca branca Bemisia argentifolii Brasília: EMBRAPA-CNPH, Cir.. Téc. 9, 11p.
Datas de Publicação
-
Publicação nesta coleção
27 Nov 2002 -
Data do Fascículo
Jun 2002
Histórico
-
Recebido
06 Jul 2001 -
Aceito
06 Jan 2002