Abstract
Many plant species from Brazilian semi-arid present arbuscular mycorrhizal fungi (AMF) in their rhizosphere. These microorganisms play a key role in the establishment, growth, survival of plants and protection against drought, pathogenic fungi and nematodes. This study presents a quantitative analysis of the AMF species associated with Mimosa tenuiflora, an important native plant of the Caatinga flora. AMF diversity, spore abundance and root colonization were estimated in seven sampling locations in the Ceará and Paraíba States, during September of 2012. There were significant differences in soil properties, spore abundance, percentage of root colonization, and AMF diversity among sites. Altogether, 18 AMF species were identified, and spores of the genera Acaulospora, Claroideoglomus, Dentiscutata, Entrophospora, Funneliformis, Gigaspora, Glomus, Racocetra, Rhizoglomus and Scutellospora were observed. AMF species diversity and their spore abundance found in M. tenuiflora rhizosphere shown that this native plant species is an important host plant to AMF communities from Brazilian semi-arid region. We concluded that: (a) during the dry period and in semi-arid conditions, there is a high spore production in M. tenuiflora root zone; and (b) soil properties, as soil pH and available phosphorous, affect AMF species diversity, thus constituting key factors for the similarity/dissimilarity of AMF communities in the M. tenuiflora root zone among sites.
Keywords
AMF community structure; AMF-plant pairing; Dryland; Glomeromycota
Introduction
The Brazilian semi-arid region is an important Caatinga ecoregion that provides the required biotic and abiotic conditions for many endemic species to inhabit; thus, having high species diversity when compared with other semi-arid regions of the World. This ecoregion covers approximately 12% of the country, and is located in the innermost region of northeastern Brazil. It is characterized by frequent dry periods, and by a unique biota, with thousands of endemic species and over 1000 species of vascular plant.11 Alves JJA, Araújo MA, Nascimento SS. Degradação da Caatinga: uma investigação ecofisiográfica. Rev Caatinga Mossoró. 2009;22:126-135.,22 Santos JC, Leal IR, Almeida-Cortez JS, Fernandes GW, Tabarelli M. Caatinga: the scientific negligence experienced by a dry tropical forest. Trop Conserv Sci. 2011;4:276-286. However, the establishment, growth and dispersal of native vegetation are often restricted by long dry periods anthropogenic activities, and exotic species introduction.11 Alves JJA, Araújo MA, Nascimento SS. Degradação da Caatinga: uma investigação ecofisiográfica. Rev Caatinga Mossoró. 2009;22:126-135.
In this context, plant mutualistic interactions with other organisms, such as fungi, might play a significant role during their establishment and growth. Mutualistic symbiosis between arbuscular mycorrhizal fungi (AMF) and plants are advantageous for both organisms involved, because the plant provides the AMF with energetic products resulting from photosynthesis, allowing AMF growth and maintenance, while AMF provides the plant with water and mineral nutrients, such as phosphorus and nitrogen.33 Smith SE, Read DJ. Mycorrhizal Symbiosis. 3rd ed. Amsterdam:Bostoh, Academic Press; 2008.,44 Hodge A, Storer K. Arbuscular mycorrhizal and nitrogen: implications for individual plants through to ecosystems. Plant Soil. 2014;386:1-19, http://dx.doi.org/10.1007/s11104-014-2162-1.
http://dx.doi.org/10.1007/s11104-014-216...
By enhancing plant's mineral nutrients uptake, AMF play a key role in the establishment and survival of native plants in this biome, promote plant growth and offer protection against drought and soil pathogens.55 Augé RM. Water relations, drought and VA mycorrhizal symbiosis. Mycorrhiza. 2001;11:3-42.
6 Cavagnaro TR, Gao L-L, Smith FA, Smith SE. Morphology of arbuscular mycorrhizas is influenced by fungal identity. New Phytol. 2001;151:469-475.
7 Koske RE, Poison WR. Are VA mycorrhizae required for sand dune stabilization? BioScience. 1984;34:420-424.-88 Rodríguez-Echeverría S, Crisóstomo JA, Nabais C, Freitas H. Belowground mutualists and the invasive ability of Acacia longifolia in coastal dunes of Portugal. Biol Invasions. 2009;11:651-661.
Mimosa tenuiflora (Willd.) Poir (Fabaceae) is a perennial native plant from northeastern Brazil. Similarly to other Fabaceae, M. tenuiflora is able to fix nitrogen because its roots form associations with bacteria from the genus Rhizobium.99 Yano-Melo AM, Trufem SFB, Maia LC. Arbuscular mycorrhizal fungi in salinized and surrounded areas at the São Francisco Submedium Valley, Brazil. Hoehnea. 2003;30:79-87. This plant species is considered to be a prolific pioneer plant,1010 LPWG (The Legume Phylogeny Working Group) Legume phylogeny and classification in the 21st century: progress, prospects and lessons for other species-rich clades. Taxon. 2013;62:217-248, doi:10.127-5/622.8
https://doi.org/10.127-5/622.8...
which is very tolerant to dry periods, fires and other major ecological disturbances.1111 Bakke IA, Bakke OA, Andrade AP, Salcedo IH. Regeneração natural da Jurema Preta em áreas sob pastejo de bovinos. Rev Caatinga Mossoró. 2006;19:228-235. During highly stress periods, its leaves start to fall, representing a mechanism for the plant to save energy.1010 LPWG (The Legume Phylogeny Working Group) Legume phylogeny and classification in the 21st century: progress, prospects and lessons for other species-rich clades. Taxon. 2013;62:217-248, doi:10.127-5/622.8
https://doi.org/10.127-5/622.8...
It develops extensive root systems, which are colonized by mutualistic organisms, as bacteria and fungi. Indeed, previous studies have shown the association between AMF and M. tenuiflora root system.99 Yano-Melo AM, Trufem SFB, Maia LC. Arbuscular mycorrhizal fungi in salinized and surrounded areas at the São Francisco Submedium Valley, Brazil. Hoehnea. 2003;30:79-87.,1212 Lima RLFA, Salcedo IH, Fraga VS. Propágulos de fungos micorrízicos arbusculares em solos deficientes em fósforo sob diferentes usos, da região semiárida no Nordeste do Brasil. Rev Bras Ciênc Solo. 2007;31:257-268.,1313 Mergulhão ACES, Oliveira JP, Burity HA, Maia LC. Potencial de infectividade de fungos micorrízicos arbusculares em áreas nativas e impactadas por mineração gesseira no semiárido brasileiro. Hoehnea. 2007;34:341-348. The primarily objective of most studies conducted have been to verify the presence of AMF, but more recently, they have focused on the AMF species diversity commonly found associated with native species of the Brazilian semi-arid region. Those studies have shown that species from the genera Archaeospora, Acaulospora, Glomus, Gigaspora and Scutellospora frequently form mutualistic symbiosis with native plant species.1414 Silva LX, Figueiredo MVB, Silva GA, et al. Fungos micorrízicos arbusculares em áreas de plantio de Leucena e Sabiá no estado de Pernambuco. Rev Árvore. 2007;31:427-435.,1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175.
Soil fertility is often reported as an important factor conditioning AMF species diversity, having effects at the community level, but also on the biological activity of these microorganisms.1616 Alguacil MM, Torrecillas E, Roldán A, Díaz G, Torres MP. Perennial plant species from semiarid gypsum soils support higher AMF diversity in roots than the annual Bromus rubens. Soil Biol Biochem. 2012;49:132-138.
17 Beauregard MS, Gauthier M-P, Hamel C, et al. Various forms of organic and inorganic P fertilizers did no negatively affect soil - and root-inhabiting AM fungi in a maize–soybean rotation system. Mycorrhiza. 2013;23:143-154.
18 Ji B, Bentivenga SP, Casper BB. Comparisons of AM fungal spore communities with the same hosts but different soil chemistries over local and geographic scales. Oecologia. 2012;168:187-197.-1919 Lekberg Y, Koide RT, Rohr JR, Aldrichwolfe L, Morton JB. Role of niche restrictions and dispersal in the composition of arbuscular mycorrhizal fungal communities. J Ecol. 2007;95:95-105. Thus, the objective of the present study was to provide a quantitative assessment of the AMF species associated with M. tenuiflora in the Brazilian xeric shrublands sampled from different locations across the Ceará and Paraíba States. To accomplish this objective, we selected seven independent sites where we sampled M. tenuiflora root zone and we hypothesized that (1) the root zone of this native plant species has high AMF spore abundance and root colonization during the dry period; and (2) soil properties are an important factor that influence the AMF community in different M. tenuiflora root zones.
Materials and methods
Site description and sampling
Sampling of the study species root zone area was performed in locations with similar importance value index2020 Magurran AE. Species abundance over time. Ecol Lett. 2007;10:347-354. (IVI, see table below) of M. tenuiflora in Ceará and Paraíba States, Brazil (Table 1). During the dry season, the average annual temperature is of 30 °C, while during the rainy season, the average annual temperature is around 23 °C. Rainfall in the greater part of this region is scanty, unpredictable and irregular.11 Alves JJA, Araújo MA, Nascimento SS. Degradação da Caatinga: uma investigação ecofisiográfica. Rev Caatinga Mossoró. 2009;22:126-135. The flowering season for M. tenuiflora starts on August in Ceará State, one month earlier than Paraíba State. The flowering starts after the rainy season (August) and during the dry season that the plant growth is very limited.
Geographic coordinates and the importance value index (IVI, %) of M. tenuiflora plants in each studied site.
In each site, we established 40 plots of 100 m2, accordingly to the methodology proposed by Grieg-smith2222 Grieg-Smith P. Quantitative Plant Ecology. Oxford: Blackwell; 1983. and Fortin and Dale.2323 Fortin M, Dale MR. Spatial Analysis: A Guide for Ecologists. Cambridge: Cambridge University Press; 2005. Before sampling M. tenuiflora root zone, the following criteria were established: (a) each plant should be larger than 3 cm in diameter near soil surface; (b) its height should vary between 1 and 3 m; and (c) no other individuals from a different plant species should occur at a distance lower than 3 m from the sampling point.2323 Fortin M, Dale MR. Spatial Analysis: A Guide for Ecologists. Cambridge: Cambridge University Press; 2005.
24 Caifa AN, Martins FR. Taxonomic identification, sampling methods, and minimum size of the tree sampled: implications and perspectives for studies in the Brazilian Atlantic Rainforest. Funct Ecosyst Commun. 2007;1:95-104.
25 Costa IR, Araújo FS. Organização comunitária de um encrave de cerrado sensu stricto no bioma Caatinga, chapada do Araripe, Barbalha, Ceará. Acta Bot Brasil. 2007;18:759-770.
26 Durigan G. Estrutura e diversidade de comunidades florestais. In: Martins SV, ed. Ecologia de Florestas Tropicais do Brasil. Vicoça, MG: Editora UFV; 2009:185–215.-2727 Daubenmire RE. Plant Communities: A Textbook of Plant Synecology. New York: Haper and Row; 1968. In total, twenty-five root zone samples per plot including soil and root fragments were collected at a depth ranging between 0 and 20 cm. Soil was kept in plastic bags and root fragments were stored in 50% ethanol until laboratory analysis. Because fungal sporulation is expected to be higher during the dry period in semiarid environments,1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175. the sampling was conducted during September of 2012.
Soil analyses
For each plot, soil and roots samples were pooled resulting in 40 samples per site. In each of these samples, the following variables were measured: pH, carbon, total nitrogen and available phosphorus. Soil pH was determined in a suspension of soil and distilled water2828 Black CA. Methods of soil analysis, Part 2. In: Black CA, ed. Agronomy Monograph No. 9. Madison, WI: American Society of Agronomy; 1965:771–1572.; soil organic carbon was estimated accordingly to the methodology described by Okalebo2929 Okalebo JR, Gathua KW, Woomer PL. Laboratory Methods of Plant and Soil Analysis: A Working Manual. Technical Bulletin No. 1 Soil Science Society East Africa; 1993.; total soil nitrogen content was estimated following the Kjeldahl method2828 Black CA. Methods of soil analysis, Part 2. In: Black CA, ed. Agronomy Monograph No. 9. Madison, WI: American Society of Agronomy; 1965:771–1572.; and available phosphorus was determined spectrophotometrically following the molybdenum blue method and using stannous chloride as the reducing agent.2828 Black CA. Methods of soil analysis, Part 2. In: Black CA, ed. Agronomy Monograph No. 9. Madison, WI: American Society of Agronomy; 1965:771–1572.
Spores assessment
Spores were extracted from field soil samples (n = 40) by wet sieving3030 Gerdemann JW, Nicolson TH. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans Br Mycol Soc. 1963;1:43-66. followed by sucrose centrifugation.3131 Jenkins WR. A rapid centrifugal flotation technique for separating nematodes from soil. Plant Dis Rep. 1964;48:692 Initially, the extracted spores were examined in aqueous solution under a dissecting microscope and separated based on morphological characteristics. Then, polyvinyl alcohol lacto-glycerol (PVLG) mixed or not with Melzer's reagent was used as permanent mounting medium.3232 Walker C, Vestberg M, Demircik F, et al. Molecular phylogeny and new taxa in the Archaeosporales (Glomeromycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycol Res. 2007;111:137-153. Species identification was based on the descriptions provided by Schenck and Perez,3333 Schenck NC, Perez Y. Manual for the Identification of VA Mycorrhizal Fungi. 3rd ed. Gainesville: Synergistic Publications; 1990. publications with descriptions of new genera and families,3434 Oehl F, Souza FA, Sieverding E. Revision of Scutellospora and description of five new genera and three new families in the arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon. 2008;106:311-360. and by consulting the international culture collection of arbuscular mycorrhizal fungi database – INVAM (http://invam.caf.wvu.edu). For the purpose of this work, we adopted the classification proposed by Oehl et al.,3535 Oehl F, Sieverding E, Palenzuela J, Ineichen K, Silva GA. Advances in Glomeromycota taxonomy and classification. IMA Fungus. 2011;2:191-199. including recently described new taxa.3636 Goto BT, Silva GA, Assis DMA, et al. Intraornatosporaceae (Gigasporales), a new family with two new genera and two new species. Mycotaxon. 2012;119:117-132.
37 Redecker D, Schüßler A, Stockinger H, et al. An evidence-based consensus for the classification or arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza. 2013-3838 Sieverding E, Silva GA, Berndt R, Oehl F. Rhizoglomus, a new genus of the Glomeraceae. Mycotaxon. 2014;129:373-386.
Mycorrhizal root colonization assessment
To assess mycorrhizal root colonization, roots were first cleared in 2% KOH for 1 h at 90 °C, and subsequently they were acidified in 1% HCl overnight. Blue ink (Parker Quink) was used as staining agent for 30 min at 60 °C, followed by a destaining treatment with lactoglycerol. The extent of mycorrhizal root colonization was estimated by using a grid-intersect method with the examination of 100 intersects under a compound microscope at 200× magnification.3939 Phillips JM, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc. 1970;55:158-160.,4040 McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990;115:495-501. Prior to microscope examinations, we decided to score hyphae as “mycorrhizal”, based on the associated presence of vesicles, arbuscules, spores, and the morphology of the mycelium. Thus, if any of the structures referred were found in one of the 100 microscope intersections per sample per study site (n = 400 intersections per study site), they were scored as “mycorrhizal”. To avoid error associated with the observer, all microscopic examinations were carried out by the same individual.
Statistical analysis
One-way ANOVA was used to compare spore abundance and root colonization among the seven locations sampled. The relationship between the AMF community structure and soil properties was examined by calculating the Pearson correlation coefficient. To explore the variability and similarities among soils and AMF communities composition among sites, principal component analysis (PCA) was used.
The following ecological indexes were calculated after AMF species identification: average species richness of AMF, diversity index4141 Shanon CE, Weaver W. The Mathematical Theory of Communication. Urbana, IL: University of Illinois Press; 1949. and dominance index4242 Simpson EH. Measurement of diversity. Nature. 1949;163:688 for each studied site. One-way ANOVA was used to compare species richness of AMF, the diversity index and the dominance index among sites. In addition, the Zhang's frequency of occurrence classification4343 Zhang Y, Gui LD, Liu RJ. Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil. 2004;261:257-263. was used to analyze (a) total spore abundance (total number of spores), (b) spore abundance of a given AMF species (number of spores of a particular AMF species), and (c) AMF species occurrence frequency (percentage of samples containing a particular AMF species). This procedure allowed AMF species to be classified as dominant (FO > 50%), most common (31 ≤ FO ≤ 50%), common (10 ≤ FO ≤ 30%) and rare (FO < 10%).4343 Zhang Y, Gui LD, Liu RJ. Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil. 2004;261:257-263. Additionally, AMF species were also classified as generalists (present in six areas), intermediate (present in two to five areas) or exclusive (restricted to only one area).4444 Stürmer SL, Siqueira JO. Species richness and spore abundance of arbuscular mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza. 2011;21:255-267.
Before statistical analysis, data was checked for normality and homogeneity of variances. One-way ANOVA and the Pearson correlation coefficients were calculated using SAS 9.1.3 Portable, while PCA analysis and the ecological index calculations were conducted using MVSP 3.1.4545 Kovach WL. MVSP – A MultiVariate Statistical Package for Windows, ver. 3.1. Pentraeth, Wales, UK: Kovach ComputingServices; 2007.
Results
Soil analysis
All soil chemical properties varied significantly among sites (Table 2). Accordingly to the PCA analysis, A4 and A6 were the most dissimilar locations (Fig. 1). A1 and A2 were more similar to each other than A3, A5 and A7. Total organic carbon and available phosphorous were the main factors contributing to the variance of most samples. PCA plot also showed a strong positive relationship between total nitrogen, soil pH and available phosphorous and 93.1% of the samples’ variation was explained by the two axes.
PCA score plot of soil properties for the seven studied sites. A1 – Algodão de Jandaíra, PB; A2 – Esperança, PB; A3 – Ibaretama, CE; A4 – Juazeirinho, PB; A5 – Monteiro, PB; A6 – Natuba, PB; A7 – Poçinhos, PB. Points represent samples from each plot by studied sites.
Spore abundance and root colonization
Sites A4 and A5 did not significantly differ in spore abundance data, showing the highest values from all studied sites, averaging 16.6 spores g-1 soil and 15.3 spores g-1 soil, respectively (Fig. 2). In the remaining five sites, we found statistically significant differences (p < 0.05) when comparing A1, A2 and A6 with A3 and A7. A7 had the smallest value of spore abundance from all studied sites (6.8 spores g-1 soil). Statistically significant differences (p < 0.05) were also found in root colonization among all sites (averages ranging between 6.3 and 36.5%). A2 had the highest root colonization, while A1 the smallest of all studied sites (Fig. 2).
AMF spore abundance, given as number of spores g−1 soil, and percentage of root colonization (mean ± SE, n = 40) in the rhizosphere of M. tenuiflora. Different letters represent statistically significant differences (p < 0.05) among sites, after one-way ANOVA and LSD test for overall comparisons.
AMF community diversity and structure
Ecological indexes significantly differed among sites (p < 0.05). AMF total richness varied between 14 and 18 species, the diversity index ranged between 1.98 and 2.64, and the values for the dominance index averaged between 0.80 and 0.91 among sites. A4 site had the highest values for all ecological indexes calculated (species richness, 18 species; diversity index, 2.64; and dominance index, 0.91; Table 3).
Field-collected spores were from two orders, Diversisporales (four families and six genera), and Glomerales (two families and four genera). Representatives belonging to four families of Diversisporales were identified: Acaulosporaceae Diversisporaceae, Entrophosporaceae, Gigasporaceae; while two families from Glomerales were recognized, Claroideoglomeraceae and Glomeraceae, representing ten genera – Acaulospora (2), Claroideoglomus (1), Dentiscutata (3), Entrophospora (1), Funneliformis (3), Gigaspora (3), Glomus (1), Racocetra (1), Rhizoglomus (2) and Scutellospora (1); in a total of 18 different AMF species. Accordingly to their occurrence frequency, AMF species were classified into two groups: rare AMF species (R, occurrence frequency lower than 10%), and common AMF species (C, occurrence frequency between 10 and 30%). The AMF species identified as rare species were: Acaulospora denticulata, Acaulospora tuberculata, Dentiscutata cerradensis, Dentiscutata erythropus, Dentiscutata heterograma, Entrophospora infrequens, Funneliformis caledonium, Funneliformis geosporum, Funneliformis mosseae, Gigaspora gigantea, Glomus multicaule, Racocetra coralloidea, Rhizoglomus aggregatum, Scutellospora calospora, Claroideoglomus etunicatum, Gigaspora albida, Gigaspora decipiens, and Rhizoglomus clarum were classified as common species. No AMF species had an occurrence frequency higher than 30% (Table 4).
Occurrence frequency (FOi ) of the AMF species identified. Values are given as percentage followed by their occurrence frequency classification between parentheses accordingly to Zhang et al. 4343 Zhang Y, Gui LD, Liu RJ. Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil. 2004;261:257-263.
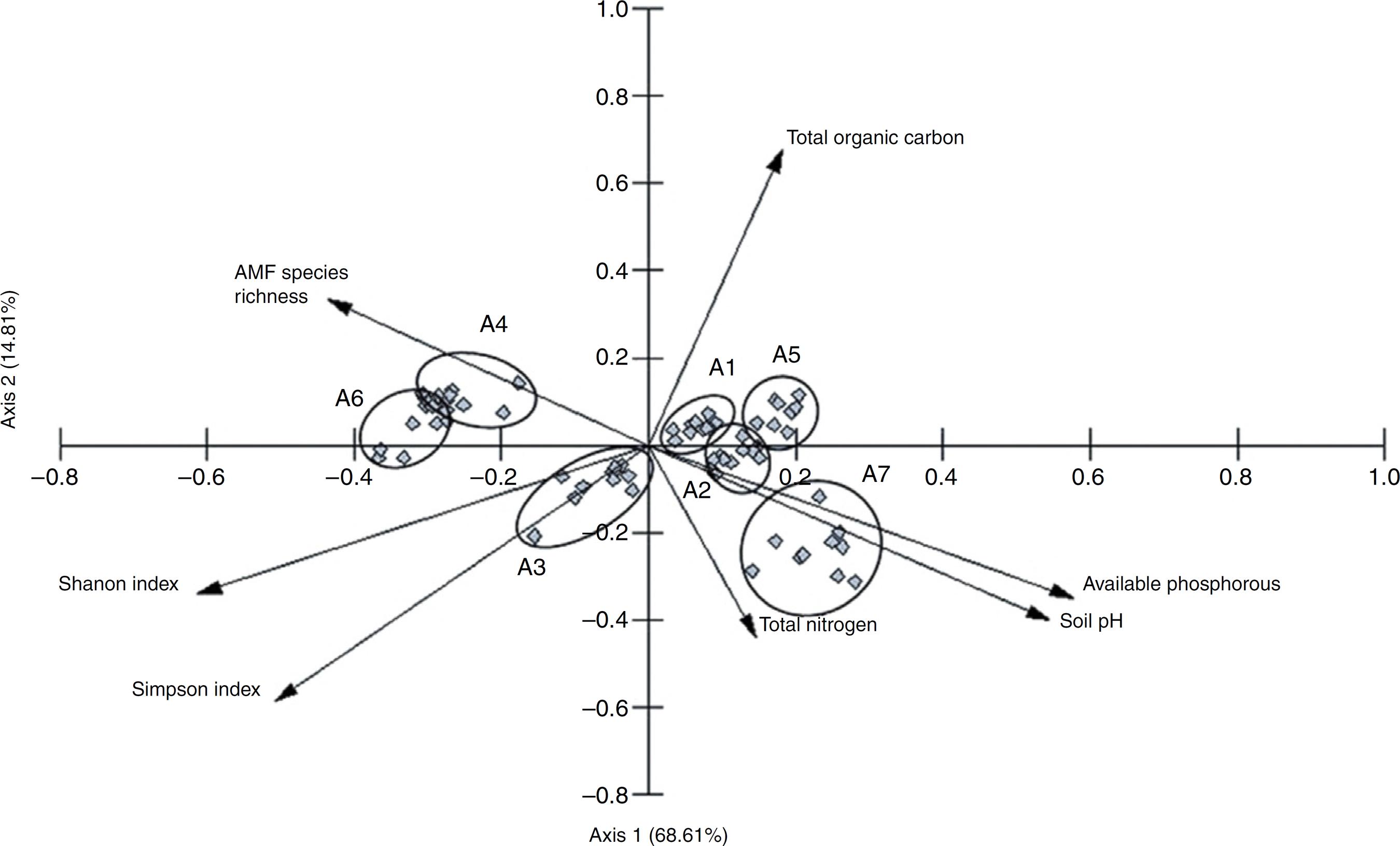
The PCA analyses showed that AMF species richness, Shanon index, Simpson index, soil pH, total organic carbon and available phosphorous were the main factors contributing to the variance of the samples (Fig. 3). The analysis also showed: (1) a strong negative relationship between total soil pH and available phosphorous with AMF species richness; and (2) a negative relationship between total organic carbon and Shanon and Simpson indexes. The two axes explained 83.42% of the samples’ variation.
Fig. 3 – PCA score plot of soil properties and AMF community structure, here represented by the ecological indexes calculated (species richness, Shanon index and Simpson index). A1 – Algodão de Jandaíra, PB; A2 – Esperança, PB; A3 – Ibaretama, CE; A4 – Juazeirinho, PB; A5 – Monteiro, PB; A6 – Natuba, PB; A7 – Poçinhos, PB. Points represent samples from each plot by studied sites.
Discussion
Our results provided evidence that AMF are common in M. tenuiflora roots and rhizosphere, supporting our hypothesis that M. tenuiflora rhizosphere has high AMF spore abundance and root colonization. However, AMF species were quantitatively variable among the studied sites. Mello et al.4646 Mello CMA, Silva IR, Pontes JS, et al. Diversidade de fungos micorrízicos arbusculares em área de Caatinga, PE, Brasil. Acta Bot Brasil. 2012;26:938-943. reported low values of spore abundance, <1 spore g-1 soil, and root colonization, <10%, in the rhizosphere of native plants from Brazilian semi-arid region, as Aspidosperma pyrifolium, Bromelia sp., Myracrodruon urundeuva, Tabebuia impetiginosa, and Zizyphus joazeiro. Contrarily, crop host plants, such as maize and sorghum, usually have high sporulation (>5 spores g-1 soil) and high root colonization values (approximately 40%).4747 Oehl F, Laczko E, Bogenrieder A, et al. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol Biochem. 2010;42:724-738. Thus, our results suggest that M. tenuiflora have a good level of promiscuity as a host plant, presenting high values of sporulation and mycorrhizal colonization independently of the studied site. In addition to the variability found in spore abundance and root colonization for all studied sites, we also observed a significant variation in the content of vesicles and arbuscules, which probably indicates different physiological and/or phenological mechanisms of the symbiosis between the host plant and the fungi among sites (Unpublished data).
The differences in spore abundance and root colonization among sites are in agreement with previous work1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175.,4848 Pereira CMR, Silva DKA, Ferreira ACA, Goto BT, Maia LC. Diversity of arbuscular mycorrhizal fungi in Atlantic forest areas under different land uses. Agric Ecosyst Environ. 2014;185:245-252. that reported variability in spore abundance during the dry season in Brazilian semiarid region for AMF species from Diversisporales order. Silva et al.1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175. concluded that the dry season is the period of greatest AMF spore production, because soil water availability plays an important role in the AMF-host plant relationship, namely in terms of the composition and functioning. Also, data available in the literature from elsewhere showed that spore abundance might vary from 0 to 13 spores g-1 soil and root colonization ranges from 27.7 to 45.7% in other semiarid and arid regions.1313 Mergulhão ACES, Oliveira JP, Burity HA, Maia LC. Potencial de infectividade de fungos micorrízicos arbusculares em áreas nativas e impactadas por mineração gesseira no semiárido brasileiro. Hoehnea. 2007;34:341-348.,4949 Carneiro RFV, Cardozo Júnior FM, Pereira LF, Araújo ASF, Silva GA. Fungos micorrízicos arbusculares como indicadores da recuperação de áreas degradadas no Nordeste do Brasil. Rev Cienc Agron. 2012;43:648-657.
Our results for total AMF species richness differ from the ones presented by Silva et al.,1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175. which found that AMF total species richness might be larger than 40 AMF species. This dissimilarity in results may be due to soil site-specific characteristics, as well as rhizosphere peculiarities that differ among sites that might determine the number of AMF species present, and consequently, the number of AMF species detected.4949 Carneiro RFV, Cardozo Júnior FM, Pereira LF, Araújo ASF, Silva GA. Fungos micorrízicos arbusculares como indicadores da recuperação de áreas degradadas no Nordeste do Brasil. Rev Cienc Agron. 2012;43:648-657. However, our results are similar to recently published data from semiarid and arid areas elsewhere, which showed total richness to vary between 3 and 32 AMF species.1616 Alguacil MM, Torrecillas E, Roldán A, Díaz G, Torres MP. Perennial plant species from semiarid gypsum soils support higher AMF diversity in roots than the annual Bromus rubens. Soil Biol Biochem. 2012;49:132-138.,5050 Wetzel K, Silva G, Matczinski U, Oehl F, Fester T. Superior differentiation of arbuscular mycorrhizal fungal communities from till and no-till plots by morphological spore identification when compared to T-RFLP. Soil Biol Biochem. 2014;72:88-96.
There was a statistically significant difference in AMF diversity and dominance among sites. A4 was the site with the greatest diversity (2.64) and the highest dominance index (0.91). Other studies carried out in the Brazilian semiarid region found similar results.1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175.,5151 Pagano MC, Zandavalli RB, Araújo FS. Biodiversity of arbuscular mycorrhizas in three vegetational types from the semiarid of Ceará State, Brazil. Appl Soil Ecol. 2013;67:37-46. As pointed out above, soil properties may have more influence than the host plant on the AMF community,1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175.,4747 Oehl F, Laczko E, Bogenrieder A, et al. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol Biochem. 2010;42:724-738.,5252 Jansa J, Erb A, Oberholzer H-R, Smilauer P, Egli S. Soil and geography are more important determinants of indigenous arbuscular mycorrhizal communities than management practices in Swiss agricultural soils. Mol Ecol. 2014;23:2118-2135 doi: 10.1111/mec.12706
https://doi.org/10.1111/mec.12706...
because soil pH, water availability, phosphorous and nitrogen contents are determinant factors in the composition and functioning of the AMF-host plant relationship. Our study provides evidence that the soil properties, such as soil pH, total nitrogen and phosphorous available, are key aspects on the AMF community structure in the Brazilian semiarid region.
Species of Acaulospora, Claroideoglomus, Dentiscutata, Funneliformis, Gigaspora, Rhizoglomus and Scutellospora were among the identified taxa. These genera are common in semiarid sites,1313 Mergulhão ACES, Oliveira JP, Burity HA, Maia LC. Potencial de infectividade de fungos micorrízicos arbusculares em áreas nativas e impactadas por mineração gesseira no semiárido brasileiro. Hoehnea. 2007;34:341-348.,1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175.,5353 Dandan Z, Zhiwei Z. Biodiversity of arbuscular mycorrhizal fungi in the hot-dry valley of the Jinsha river, southwest China. Appl Soil Ecol. 2007;37:118-128. and acid soils with low phosphorous availability.5454 Ramos AC, Façanha AR, Feijó JA. Proton (H+) flux signature for the presymbiotic development of the arbuscular mycorrhizal fungi. New Phytologist. 2008;178:177-188. Previous studies have shown the presence of these AMF species associated with M. tenuiflora.99 Yano-Melo AM, Trufem SFB, Maia LC. Arbuscular mycorrhizal fungi in salinized and surrounded areas at the São Francisco Submedium Valley, Brazil. Hoehnea. 2003;30:79-87.,1212 Lima RLFA, Salcedo IH, Fraga VS. Propágulos de fungos micorrízicos arbusculares em solos deficientes em fósforo sob diferentes usos, da região semiárida no Nordeste do Brasil. Rev Bras Ciênc Solo. 2007;31:257-268.,1313 Mergulhão ACES, Oliveira JP, Burity HA, Maia LC. Potencial de infectividade de fungos micorrízicos arbusculares em áreas nativas e impactadas por mineração gesseira no semiárido brasileiro. Hoehnea. 2007;34:341-348. Most studies have shown a certain constancy in the AMF genera associated with this plant species, like Acaulospora, Archaeospora, Gigaspora, Glomus, and Scutellospora, 1414 Silva LX, Figueiredo MVB, Silva GA, et al. Fungos micorrízicos arbusculares em áreas de plantio de Leucena e Sabiá no estado de Pernambuco. Rev Árvore. 2007;31:427-435.,1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175. but there are no reports of dominant AMF genus in the M. tenuiflora rhizosphere.4646 Mello CMA, Silva IR, Pontes JS, et al. Diversidade de fungos micorrízicos arbusculares em área de Caatinga, PE, Brasil. Acta Bot Brasil. 2012;26:938-943. Our results also support the lack of dominance of a given AMF genus or species associated with M. tenuiflora rhizosphere. Moreover, the high abundance of representatives from the Order Gigasporales found in our study, might give some support to the hypothesis proposed by Goto et al.3636 Goto BT, Silva GA, Assis DMA, et al. Intraornatosporaceae (Gigasporales), a new family with two new genera and two new species. Mycotaxon. 2012;119:117-132. and Marinho et al.,5555 Marinho F, Silva GA, Ferreira ACA, et al. Bulbospora minima, a new species in the Gigasporales from semi-arid Northeast Brazil. Sydowia. 2014;66:313-323. which points out that Brazil is the diversification and dispersion center of species from Gigasporales.
Soil properties differed among all studied sites. These results support our hypothesis that soil properties are important factors influencing the AMF community in the M. tenuiflora rhizosphere. Accordingly to Oehl et al.3535 Oehl F, Sieverding E, Palenzuela J, Ineichen K, Silva GA. Advances in Glomeromycota taxonomy and classification. IMA Fungus. 2011;2:191-199. and Carneiro et al.,4949 Carneiro RFV, Cardozo Júnior FM, Pereira LF, Araújo ASF, Silva GA. Fungos micorrízicos arbusculares como indicadores da recuperação de áreas degradadas no Nordeste do Brasil. Rev Cienc Agron. 2012;43:648-657. soil pH, available phosphorous, and other soil properties can change AMF community composition, increasing the frequency of mutualists from genera Acaulospora, Funneliformis, Gigaspora and Scutellospora. There was a negative correlation between soil pH, total nitrogen and phosphorous availability and the different ecological indices calculated for each site (Fig. 3). Our results are in agreement with previous work carried out in 16 locations across Europe,4747 Oehl F, Laczko E, Bogenrieder A, et al. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol Biochem. 2010;42:724-738. which found that different soil types contained different AMF species. In Switzerland, a study analyzing 154 different agricultural soil samples reported that the abundance of some AMF taxa was also related with soil variables.5252 Jansa J, Erb A, Oberholzer H-R, Smilauer P, Egli S. Soil and geography are more important determinants of indigenous arbuscular mycorrhizal communities than management practices in Swiss agricultural soils. Mol Ecol. 2014;23:2118-2135 doi: 10.1111/mec.12706
https://doi.org/10.1111/mec.12706...
More recently, a study performed along an environmental gradient in the Brazilian semiarid region found a strong correlation between soil attributes and AMF community composition, concluding that soil properties are key factors determining AMF community structure in a local scale, even though sites are geographically close to each other's.1515 Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol. 2014;84:166-175.
In conclusion, our results showed that (1) AMF species exhibit high sporulation during the dry season, (2) and that M. tenuiflora is a common host plant, which points out its importance in the biome it inhabits and its potential use in reforestation programs in the Caatinga degraded areas. However, further research is needed to unravel the effect of AMF inoculation in the native plants, M. tenuiflora in this particular case, growing in different soil conditions and with different AMF communities before establish habitat restoration measures. Despite no dominant AMF species was found in M. tenuiflora rhizosphere, rare and common AMF species accordingly to Zhang's classification4343 Zhang Y, Gui LD, Liu RJ. Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil. 2004;261:257-263. were described. Based on these results, one can assume that this native plant species does not benefit any AMF particular genus and highlights its potential in AMF communities’ restoration in degraded soils. Further work is needed to understand whether this diverse AMF community has a positive differential effect in increasing native plant growth, resistance or tolerance to root pathogens in the Brazilian semiarid region.5656 Rodríguez-Echeverría S, Gera Hol WH, Freitas H, Eason WR, Cook R. Arbuscular mycorrhizal fungi of Ammophila arenaria (L.) Link: spore abundance and root colonization in six locations of the European coast. Eur J Soil Biol. 2008;44:30-36.,5757 Zhang Q, Yang R, Tang J, et al. Positive feedback between mycorrhizal fungi and plants influences plant invasion success and resistance to invasion. PLoS ONE. 2010;5:e12380
Acknowledgements
Special thanks to Joana Costa for valuable discussions and checking of English grammar. Two anonymous reviewers provide helpful comments, which greatly improved a previous version of the manuscript.
References
-
1Alves JJA, Araújo MA, Nascimento SS. Degradação da Caatinga: uma investigação ecofisiográfica. Rev Caatinga Mossoró 2009;22:126-135.
-
2Santos JC, Leal IR, Almeida-Cortez JS, Fernandes GW, Tabarelli M. Caatinga: the scientific negligence experienced by a dry tropical forest. Trop Conserv Sci 2011;4:276-286.
-
3Smith SE, Read DJ. Mycorrhizal Symbiosis 3rd ed. Amsterdam:Bostoh, Academic Press; 2008.
-
4Hodge A, Storer K. Arbuscular mycorrhizal and nitrogen: implications for individual plants through to ecosystems. Plant Soil 2014;386:1-19, http://dx.doi.org/10.1007/s11104-014-2162-1
» http://dx.doi.org/10.1007/s11104-014-2162-1 -
5Augé RM. Water relations, drought and VA mycorrhizal symbiosis. Mycorrhiza 2001;11:3-42.
-
6Cavagnaro TR, Gao L-L, Smith FA, Smith SE. Morphology of arbuscular mycorrhizas is influenced by fungal identity. New Phytol 2001;151:469-475.
-
7Koske RE, Poison WR. Are VA mycorrhizae required for sand dune stabilization? BioScience 1984;34:420-424.
-
8Rodríguez-Echeverría S, Crisóstomo JA, Nabais C, Freitas H. Belowground mutualists and the invasive ability of Acacia longifolia in coastal dunes of Portugal. Biol Invasions 2009;11:651-661.
-
9Yano-Melo AM, Trufem SFB, Maia LC. Arbuscular mycorrhizal fungi in salinized and surrounded areas at the São Francisco Submedium Valley, Brazil. Hoehnea 2003;30:79-87.
-
10LPWG (The Legume Phylogeny Working Group) Legume phylogeny and classification in the 21st century: progress, prospects and lessons for other species-rich clades. Taxon 2013;62:217-248, doi:10.127-5/622.8
» https://doi.org/10.127-5/622.8 -
11Bakke IA, Bakke OA, Andrade AP, Salcedo IH. Regeneração natural da Jurema Preta em áreas sob pastejo de bovinos. Rev Caatinga Mossoró 2006;19:228-235.
-
12Lima RLFA, Salcedo IH, Fraga VS. Propágulos de fungos micorrízicos arbusculares em solos deficientes em fósforo sob diferentes usos, da região semiárida no Nordeste do Brasil. Rev Bras Ciênc Solo 2007;31:257-268.
-
13Mergulhão ACES, Oliveira JP, Burity HA, Maia LC. Potencial de infectividade de fungos micorrízicos arbusculares em áreas nativas e impactadas por mineração gesseira no semiárido brasileiro. Hoehnea 2007;34:341-348.
-
14Silva LX, Figueiredo MVB, Silva GA, et al. Fungos micorrízicos arbusculares em áreas de plantio de Leucena e Sabiá no estado de Pernambuco. Rev Árvore 2007;31:427-435.
-
15Silva IRS, Mello CMA, Ferreira Neto RA, et al. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semiarid. Appl Soil Ecol 2014;84:166-175.
-
16Alguacil MM, Torrecillas E, Roldán A, Díaz G, Torres MP. Perennial plant species from semiarid gypsum soils support higher AMF diversity in roots than the annual Bromus rubens. Soil Biol Biochem 2012;49:132-138.
-
17Beauregard MS, Gauthier M-P, Hamel C, et al. Various forms of organic and inorganic P fertilizers did no negatively affect soil - and root-inhabiting AM fungi in a maize–soybean rotation system. Mycorrhiza 2013;23:143-154.
-
18Ji B, Bentivenga SP, Casper BB. Comparisons of AM fungal spore communities with the same hosts but different soil chemistries over local and geographic scales. Oecologia 2012;168:187-197.
-
19Lekberg Y, Koide RT, Rohr JR, Aldrichwolfe L, Morton JB. Role of niche restrictions and dispersal in the composition of arbuscular mycorrhizal fungal communities. J Ecol 2007;95:95-105.
-
20Magurran AE. Species abundance over time. Ecol Lett 2007;10:347-354.
-
21WRB (IUSS WORKING GROUP). World Reference Base for Soil. World Soil Resources Reports. n. 103 Rome: FAO; 2006.
-
22Grieg-Smith P. Quantitative Plant Ecology Oxford: Blackwell; 1983.
-
23Fortin M, Dale MR. Spatial Analysis: A Guide for Ecologists Cambridge: Cambridge University Press; 2005.
-
24Caifa AN, Martins FR. Taxonomic identification, sampling methods, and minimum size of the tree sampled: implications and perspectives for studies in the Brazilian Atlantic Rainforest. Funct Ecosyst Commun 2007;1:95-104.
-
25Costa IR, Araújo FS. Organização comunitária de um encrave de cerrado sensu stricto no bioma Caatinga, chapada do Araripe, Barbalha, Ceará. Acta Bot Brasil 2007;18:759-770.
-
26Durigan G. Estrutura e diversidade de comunidades florestais. In: Martins SV, ed. Ecologia de Florestas Tropicais do Brasil Vicoça, MG: Editora UFV; 2009:185–215.
-
27Daubenmire RE. Plant Communities: A Textbook of Plant Synecology New York: Haper and Row; 1968.
-
28Black CA. Methods of soil analysis, Part 2. In: Black CA, ed. Agronomy Monograph No. 9 Madison, WI: American Society of Agronomy; 1965:771–1572.
-
29Okalebo JR, Gathua KW, Woomer PL. Laboratory Methods of Plant and Soil Analysis: A Working Manual Technical Bulletin No. 1 Soil Science Society East Africa; 1993.
-
30Gerdemann JW, Nicolson TH. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans Br Mycol Soc 1963;1:43-66.
-
31Jenkins WR. A rapid centrifugal flotation technique for separating nematodes from soil. Plant Dis Rep 1964;48:692
-
32Walker C, Vestberg M, Demircik F, et al. Molecular phylogeny and new taxa in the Archaeosporales (Glomeromycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycol Res 2007;111:137-153.
-
33Schenck NC, Perez Y. Manual for the Identification of VA Mycorrhizal Fungi 3rd ed. Gainesville: Synergistic Publications; 1990.
-
34Oehl F, Souza FA, Sieverding E. Revision of Scutellospora and description of five new genera and three new families in the arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon 2008;106:311-360.
-
35Oehl F, Sieverding E, Palenzuela J, Ineichen K, Silva GA. Advances in Glomeromycota taxonomy and classification. IMA Fungus 2011;2:191-199.
-
36Goto BT, Silva GA, Assis DMA, et al. Intraornatosporaceae (Gigasporales), a new family with two new genera and two new species. Mycotaxon 2012;119:117-132.
-
37Redecker D, Schüßler A, Stockinger H, et al. An evidence-based consensus for the classification or arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013
-
38Sieverding E, Silva GA, Berndt R, Oehl F. Rhizoglomus, a new genus of the Glomeraceae. Mycotaxon 2014;129:373-386.
-
39Phillips JM, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc 1970;55:158-160.
-
40McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol 1990;115:495-501.
-
41Shanon CE, Weaver W. The Mathematical Theory of Communication Urbana, IL: University of Illinois Press; 1949.
-
42Simpson EH. Measurement of diversity. Nature 1949;163:688
-
43Zhang Y, Gui LD, Liu RJ. Survey of arbuscular mycorrhizal fungi in deforested and natural forest land in the subtropical region of Dujiangyan, southwest China. Plant Soil 2004;261:257-263.
-
44Stürmer SL, Siqueira JO. Species richness and spore abundance of arbuscular mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza 2011;21:255-267.
-
45Kovach WL. MVSP – A MultiVariate Statistical Package for Windows, ver. 3.1. Pentraeth, Wales, UK: Kovach ComputingServices; 2007.
-
46Mello CMA, Silva IR, Pontes JS, et al. Diversidade de fungos micorrízicos arbusculares em área de Caatinga, PE, Brasil. Acta Bot Brasil 2012;26:938-943.
-
47Oehl F, Laczko E, Bogenrieder A, et al. Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol Biochem 2010;42:724-738.
-
48Pereira CMR, Silva DKA, Ferreira ACA, Goto BT, Maia LC. Diversity of arbuscular mycorrhizal fungi in Atlantic forest areas under different land uses. Agric Ecosyst Environ 2014;185:245-252.
-
49Carneiro RFV, Cardozo Júnior FM, Pereira LF, Araújo ASF, Silva GA. Fungos micorrízicos arbusculares como indicadores da recuperação de áreas degradadas no Nordeste do Brasil. Rev Cienc Agron 2012;43:648-657.
-
50Wetzel K, Silva G, Matczinski U, Oehl F, Fester T. Superior differentiation of arbuscular mycorrhizal fungal communities from till and no-till plots by morphological spore identification when compared to T-RFLP. Soil Biol Biochem 2014;72:88-96.
-
51Pagano MC, Zandavalli RB, Araújo FS. Biodiversity of arbuscular mycorrhizas in three vegetational types from the semiarid of Ceará State, Brazil. Appl Soil Ecol 2013;67:37-46.
-
52Jansa J, Erb A, Oberholzer H-R, Smilauer P, Egli S. Soil and geography are more important determinants of indigenous arbuscular mycorrhizal communities than management practices in Swiss agricultural soils. Mol Ecol. 2014;23:2118-2135 doi: 10.1111/mec.12706
» https://doi.org/10.1111/mec.12706 -
53Dandan Z, Zhiwei Z. Biodiversity of arbuscular mycorrhizal fungi in the hot-dry valley of the Jinsha river, southwest China. Appl Soil Ecol 2007;37:118-128.
-
54Ramos AC, Façanha AR, Feijó JA. Proton (H+) flux signature for the presymbiotic development of the arbuscular mycorrhizal fungi. New Phytologist 2008;178:177-188.
-
55Marinho F, Silva GA, Ferreira ACA, et al. Bulbospora minima, a new species in the Gigasporales from semi-arid Northeast Brazil. Sydowia 2014;66:313-323.
-
56Rodríguez-Echeverría S, Gera Hol WH, Freitas H, Eason WR, Cook R. Arbuscular mycorrhizal fungi of Ammophila arenaria (L.) Link: spore abundance and root colonization in six locations of the European coast. Eur J Soil Biol 2008;44:30-36.
-
57Zhang Q, Yang R, Tang J, et al. Positive feedback between mycorrhizal fungi and plants influences plant invasion success and resistance to invasion. PLoS ONE 2010;5:e12380
Publication Dates
-
Publication in this collection
Apr-Jun 2016
History
-
Received
12 Apr 2015 -
Accepted
21 Sept 2015