ABSTRACT
Earthworms are known to improve soil fertility by altering the physicochemical properties of soil. However, the changes in properties of biogenic aggregates (produced by earthworms) in regions with different vegetation covers and soils of Inceptisol toposequence in the semi-arid region have not been studied. The objective of this work was to determine the variations of the physical, chemical, and mineralogical attributes and organic fractions of biogenic aggregates of earthworms under different vegetation covers and compare them to an Inceptisol toposequence in the semi-arid region of Pernambuco State, Brazil. For this study, four Inceptisol soil profiles were selected. The sampled points were P1-upper; P2-middle third; P3-middle third; and P4-lower slope, and the soils were collected at a depth of 0-0.20 m. The biogenic aggregates were found in the P3 samples collected from three areas with different vegetation covers: an area with mesquite tree vegetation (Prosopis juliflora [Sw] D.C.); an area with neem tree vegetation (Azadirachta indica A. Juss); and an area with forage palm and mesquite tree vegetation (Opuntia sp. and Nopalea sp.). Results indicate that the biogenic aggregates were in the form of irregular blocks with a size greater than 40 mm. As compared to Inceptisol samples, these aggregates tended to select particles of smaller diameter and exhibited higher concentrations of clay minerals. The chemical and organic fraction analysis revealed that the aggregates exhibited higher Ca2+, total organic carbon (TOC), labile and humic substances, regardless of the vegetation cover. The mineralogical analysis revealed that the action of earthworms did not change the quantity or type of clay minerals.
Index terms:
Coprolites; labile carbon; soil fertility; secondary minerals; humic substance
RESUMO
As minhocas são conhecidas por melhorar a fertilidade do solo, alterando suas propriedades físico-químicas. No entanto, mudanças nas propriedades dos agregados biogênicos (produzidos por minhocas) em regiões com diferentes coberturas vegetais e, solos de topossequência de Cambissolo, na região semiárida, ainda não foram estudadas. O objetivo deste trabalho foi determinar as variações dos atributos físicos, químicos, mineralógicos e frações orgânicas de agregados biogênicos de minhocas sob diferentes coberturas vegetais e compará-los a uma toposseqüência de um Cambissolo na região semiárida do Estado de Pernambuco, Brasil. Para este estudo, quatro perfis de Cambissolo foram selecionados. Os pontos amostrados foram: P1-superior; P2 - terço médio; P3 - terço médio; e P4 - declive inferior, e os solos foram coletados na profundidade de 0-0,20 m. As amostras dos agregados biogênicos foram encontradas no terço médio (P3) e coletadas em três áreas com diferentes coberturas vegetais: uma área com vegetação de algaroba (Prosopis juliflora [Sw] D.C.); uma área com vegetação arbórea de nim (Azadirachta indica A. Juss); e uma área com palmeiras forrageiras e vegetação de algaroba (Opuntia sp. e Nopalea sp.). Os resultados indicam que os agregados biogênicos estavam na forma de blocos irregulares com um tamanho maior que 40 mm. Em comparação com as amostras de Cambissolo, esses agregados tenderam a selecionar partículas de solo de menor diâmetro e exibiram maiores concentrações de minerais de argila. A análise química e da fração orgânica revelou que os agregados exibiram maior Ca2+, carbono orgânico total (COT), substâncias lábeis e húmicas, independentemente da cobertura vegetal. A análise mineralógica revelou que a ação das minhocas não alterou a quantidade ou tipo de argilominerais.
Termos para indexação:
Coprólitos; carbono lábil; fertilidade do solo; minerais secundários; substância húmica.
INTRODUCTION
The soil biota is known to improve soil productivity by bringing about various modifications in the soil, mainly related to the maintenance of fertility and its natural quality. Earthworms form a major part of the soil fauna. Their activity is mostly considered beneficial as they promote changes in the chemical and physical characteristics of soil through biogenic structures (Fusaro et al., 2018FUSARO, S. et al. Soil Biological Quality Index based on earthworms (QBS-e). A new way to use earthworms as bioindicators in agroecosystems. Ecological Indicators, 93:1276-1292, 2018. ; Xiang et al., 2018XIANG, H. et al. In situ earthworm breeding to improve soil aggregation, chemical properties, and enzyme activity in Papayas. Sustainability , 10(4):1193-1206, 2018. ; Mergen Junior et al., 2019aMERGEN JUNIOR, C. A. et al. Atributos químicos em agregados biogênicos e fisiogênicos de solo submetido à aplicação com dejetos suínos. Revista Brasileira de Ciências Agrárias, 14(1):1-8, 2019a., 2019bMERGEN JUNIOR, C. A. et al. Caracterização física de agregados do solo submetido a 10 anos de aplicação de dejetos suínos. Revista Brasileira de Ciências Agrárias , 36(1):79-92, 2019b. ; Melo et al., 2020MELO, M. F. et al. Vermicompostagem: Conversão de resíduos orgânicos em benefícios para solo e plantas. In: ANDRADE, D. F. (Ed.). Tópicos em Ciências Agrárias. Belo Horizonte, Minas Gerais: Poisson, v. 6, p. 35-46, 2020. ; Teshajonova; Valihanov, 2020TESHAJONOVA, M. M.; VALIHANOV, A. O. Some information about earthworms and their biology. Academicia: An International Multidisciplinary Research Journal, 10(12):1171-1176, 2020. ). Among the different types of earthworms, geophagous earthworms often have the ability to move both horizontally and vertically; this activity contributes to changes in physical attributes of soil, such as an increase in porosity, aeration, aggregation, water retention capacity, and reduction of erosive processes (Jouquet; Blanchart; Capowiez, 2014JOUQUET, P.; BLANCHART, E.; CAPOWIEZ, Y. Utilization of earthworms and termites for the restoration of ecosystem functioning. Applied Soil Ecology, 73:34-40, 2014.; Hallam; Hodson, 2020HALLAM, J.; HODSON, M. E. Impact of different earthworm ecotypes on water stable aggregates and soil water holding capacity. Biology and Fertility of Soils, 56:607-617, 2020. ).
In addition, the intense movement of earthworms also improves the chemical properties and fertility of the soil by promoting the incorporation of organic material in the entire profile of soil (Brown; Dominguez, 2010BROWN, G. G.; DOMÍNGUEZ, J. Uso das minhocas como bioindicadoras ambientais: princípios e práticas - o 3° Encontro Latino Americano de Ecologia e Taxonomia de Oligoquetas (ELAETAO3). Acta Zoológica Mexicana, 26(2):1-18, 2010.). Apart from this, the differentiated selection of food (organic and/or mineral compounds in different proportions) and the biochemical reactions due to digestion in earthworms spatially alter the distribution of organic and mineral particles, thereby increasing the concentration and availability of nutrients in the soil (Véle et al., 2011VÉLE, A. et al. Local and landscape drivers of ant and carabid beetle communities during spruce forest succession. European Journal of Soil Biology , 47(6):349-356, 2011. ; Jouquet; Blanchart; Capowiez, 2014JOUQUET, P.; BLANCHART, E.; CAPOWIEZ, Y. Utilization of earthworms and termites for the restoration of ecosystem functioning. Applied Soil Ecology, 73:34-40, 2014.; Teshajonova; Valihanov, 2020TESHAJONOVA, M. M.; VALIHANOV, A. O. Some information about earthworms and their biology. Academicia: An International Multidisciplinary Research Journal, 10(12):1171-1176, 2020. ).
The earthworms can change the soil organic matter (SOM) by selectively foraging organic particles and then mixing it with mucus and microorganisms present in their intestines, thereby accelerating or delaying the decomposition and mineralization dynamics (Bityutskii; Maiorov; Orlova, 2012BITYUTSKII, N. P.; MAIOROV, E. I.; ORLOVA, N. E. The priming effects induced by earthworm mucus on mineralization and humification of plant residues. European Journal of Soil Biology, 50:1-6, 2012. ). Rodrigues Pinto et al. (2021RODRIGUES PINTO, L. A. S. et al. Physicogenic and biogenic aggregates under different management systems in the Cerrado region, Brazil. Revista Brasileira de Ciência do Solo , 45:e0200114, 2021. ) observed that the biogenic aggregates produced by earthworms changed the SOM dynamics. Of all organic matter ingested by earthworms, a part is assimilated, and another part is added to their excreta, which are known as coprolites or biogenic structures or biogenic aggregates. These are deposited on the soil surface or subsurface.
These biogenic aggregates function as carbon sequestration and immobilization structures and can accelerate nutrient cycling (Lavelle; Rangel; Kanyonyo, 1983LAVELLE, P.; RANGEL, P.; KANYONYO, J. Mucus production by two species of tropical earthworms: Millsonia lamtoiana (Megascolecidae) and Pontoscolex corethrurus (Glossoscolecidae). In: LEBRUM, P. et al. (Eds.). New trends in soil biology. Louvain-la-Neuve: Dieu-Brichard, p.405-410, 1983. ; Rossi et al., 2006ROSSI, J. P. et al. Soil properties inside earthworm patches and gaps in a tropical grassland. European Journal of Soil Biology , 42(1):S284-S288, 2006. ; Rodrigues Pinto et al., 2021). However, the composition of biogenic aggregates varies (Fernandes et al., 2017FERNANDES, J. C. F. et al. Caracterização de agregados biogênicos, intermediários e fisiogênicos em áreas sob domínio da Mata Atlântica Brasileira. Revista Caatinga, 30(1):59-67, 2017. ; Teshajonova; Valihanov, 2020TESHAJONOVA, M. M.; VALIHANOV, A. O. Some information about earthworms and their biology. Academicia: An International Multidisciplinary Research Journal, 10(12):1171-1176, 2020. ; Rodrigues Pinto et al., 2021RODRIGUES PINTO, L. A. S. et al. Physicogenic and biogenic aggregates under different management systems in the Cerrado region, Brazil. Revista Brasileira de Ciência do Solo , 45:e0200114, 2021. ), which encourages the analysis of these aggregates in different vegetation coverings in semi-arid regions. Thus, the hypothesis of this study is that the vegetation of an area affects the physical and chemical composition and organic matter fractions of biogenic aggregates and soil. Furthermore, it has been known that earthworms interfere with the mineralogical composition of the ingested soil. Regarding SOM, most studies focus on the determination of total organic carbon (TOC); however, in the short term, small changes in TOC do not occur. Therefore, it is important to study more labile (labile carbon [LC]) and recalcitrant fractions (humic substances) of SOM.
Research indicates that there is an interdependent relationship among soil, relief (the shape of the landscape), and vegetation. The study of toposequence has been utilized to understand the pedogenesis and the current behavior of soil, mainly for establishing relationships between soil attributes and relief (Silva et al., 2007SILVA, A. C. et al. Relações entre matéria orgânica do solo e declividade de vertentes em toposseqüência de Latossolos do Sul de Minas Gerais. Revista Brasileira de Ciência do Solo , 31(5):1059-1068, 2007. ; Meireles et al., 2012MEIRELES, H. T. et al. Relações solo-paisagem em topossequência de origem basáltica. Pesquisa Agropecuária Tropical , 42(2):129-136, 2012. ; Gama-Rodrigues et al., 2018GAMA-RODRIGUES, E. F. et al. Atributos físicos, químicos e microbiológicos dos solos sob diferentes usos em topossequências no Noroeste Fluminense. Revista Brasileira de Ciências Agrarias, 13(3):e5543, 2018. ). However, research on biogenic aggregates of earthworms has mainly focused on the places where they are found and has not studied the impact of the process as a whole. Therefore, this change in the soil due to the presence of macrofauna needs to be compared with a soil toposequence. Such a knowledge gap would lead to an underestimation of the effect of these edaphic organisms on organic and chemical fractions and the mineralogy of soil. Research focusing on this aspect would provide information that can support intervention measures for the preservation of soil in the semi-arid region.
Thus, the objective of this work was to determine the variations of the physical, chemical, and mineralogical attributes and organic fractions of biogenic aggregates of earthworms found in different vegetation covers and to compare them to an Inceptisol toposequence in the semi-arid region of Pernambuco State, Brazil.
MATERIAL AND METHODS
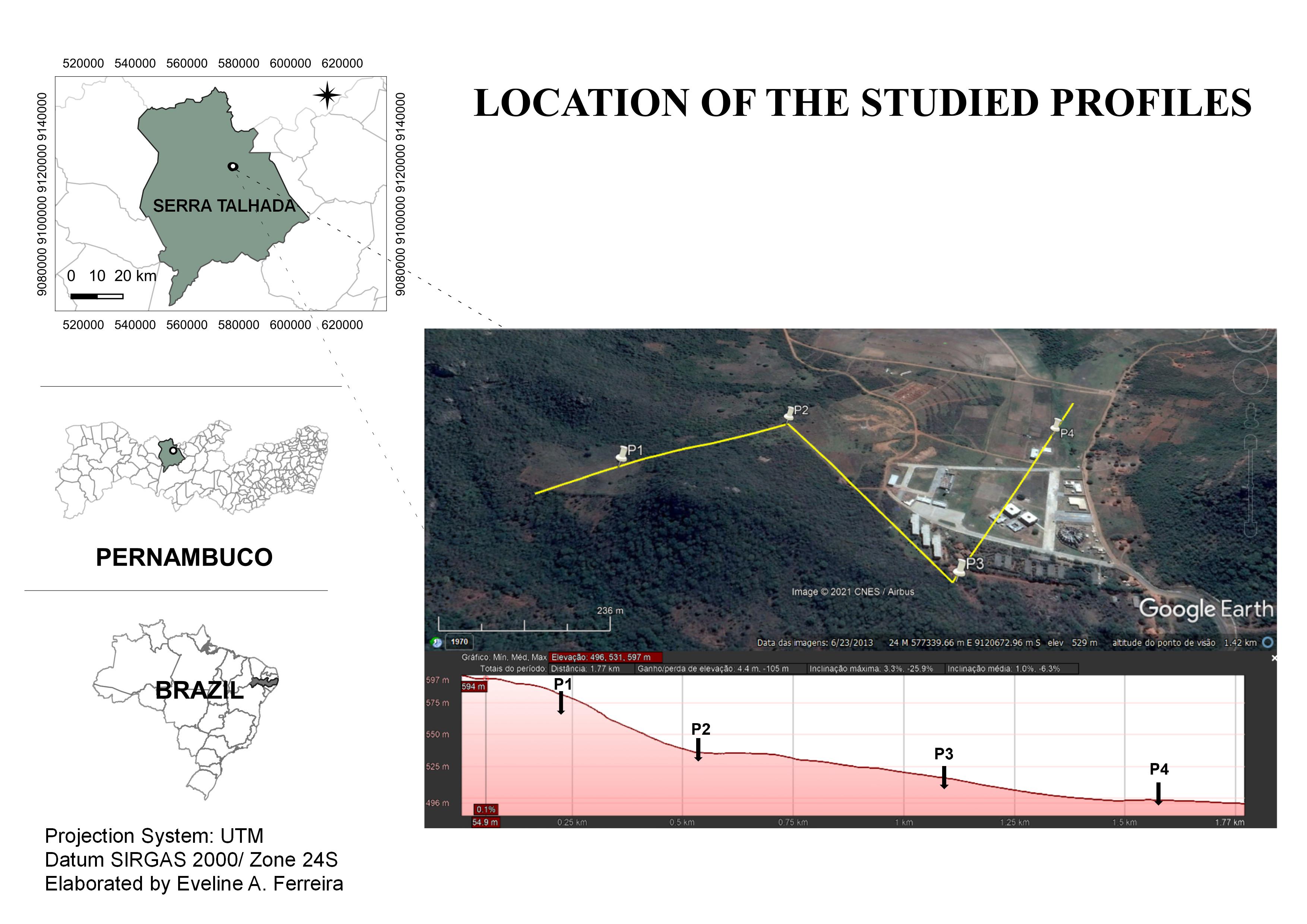
The study was carried out with biogenic aggregates and an Inceptisol toposequence located in the semi-arid region of the Pernambuco state, Brazil. The location map of the study area and the soil toposequence are shown in Figure 1.

Location map of the study area and the soil toposequence in the semi-arid region of the Pernambuco state, Brazil.
According to Köppen climate classification, the climate of this semi-arid region is classified as BSh, with an annual mean temperature of 26.5 °C. Rainfall of less than 650 mm concentrated from February to June is observed, with predominant Caatinga vegetation (Alvares et al., 2013ALVARES, C. A. et al. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22(6):711-728, 2013. ). The soil was described morphologically according to Santos et al. (2013SANTOS, R. D. et al. Manual de descrição e coleta de solo no campo. 6.ed. rev. e ampl. Viçosa: Sociedade Brasileira de Ciência do Solo, 2013. 100p.) and classified as Cambissolo Háplico Ta Eutrófico according to Sistema Brasileiro de Classificação do Solo-SiBCS (Empresa Brasileira de Pesquisa Agropecuária - Embrapa, 2018EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Sistema Brasileiro de classificação de solos. 5. ed. rev. e ampl. − Brasília, DF: Embrapa, 2018. 356p.). According to World Reference Base For Soil Resources (WRB), this soil is classified as Cambisols (Eutric Cambisols) (IUSS Working Group-WRB, 2015IUSS WORKING GROUP WRB. 2015. World reference base for soil resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. Rome, Italy: FAO, 2015. 203p. ), and in the United States of America, it is named Inceptisols (Typic Haplustept) (Soil Taxonomy, 1999SOIL TAXONOMY. A Basic system of soil classification for making and interpreting soil surveys. Washington, DC: United States Department of Agriculture (USDA), 1999. 886p. ).
For this study, four profiles of Inceptisol toposequence were selected. Horizon A samples were collected at the points P1-upper; P2-middle third; P3-middle third; and P4-lower slope. Deformed samples were collected from the horizons in the form of a trench of 1.5 × 1.5 × 1.20 meters (width, length, and depth, respectively). For analyses, five composite samples (repeats) from five sub-samples were taken for each profile at a depth of 0-0.20 m.
Samples of biogenic aggregates of earthworms were located in the P3-A point, and six aggregates were randomly collected from the soil surface of each area and classified according to size and morphology defined by Melo Júnior (2012MELO JÚNIOR, H. B. et al. Efeito da ação decompositora da minhoca californiana (Lumbricus rubellus) na composição química de um fertilizante organomineral. Bioscience Journal, 28(1):170-178, 2012. ). No biogenic aggregates were found in the other soil profiles. These biogenic aggregates were collected from three distinct areas with different vegetation types: BA1-biogenic aggregates in an area with mesquite tree vegetation (Prosopis juliflora [Sw] D.C.); BA2-biogenic aggregates in an area with neem tree vegetation (Azadirachta indica A. Juss); and BA3-biogenic aggregates in an area with forage palm and mesquite tree vegetation (Opuntia sp. and Nopalea sp.).
All samples (soils and aggregates) were air-dried, ground, and passed through a 2-mm mesh sieve to obtain “air-dried fine soil” (ADFS). Subsequently, granulometric analyses were carried out by collecting the suspension (silt + clay), and the sand was separated by sieving according to the methodology of Ruiz (2005RUIZ, H. A. Incremento da exatidão da análise granulométrica do solo por meio da coleta da suspensão (silte + argila). Revista Brasileira de Ciência do Solo , 29(5):297-300, 2005. ). All chemical analyses were performed by following the methodology of Donagema et al. (2011DONAGEMA, G. K. et al. Manual de métodos de análise de solos. 2ª ed. rev. Rio de Janeiro: Embrapa Solos, 2011. 230p. (Embrapa Solos. Documentos, 132). ). The extraction of available phosphorous (P), sodium (Na), and potassium (K) was performed by the Mehlich-1 method; the determination of P levels was performed by colorimetry, and the determination of Na+ and K+ was performed by flame photometry. The extraction of exchangeable cations, such as Ca2+ and Mg2+, was carried out with KCl at a concentration of 1 mol L-1, and the determination of their contents was performed by atomic absorption spectrophotometry. Soil pH (H2O) (1:2.5) and the electrical conductivity (EC) of the saturated paste extract (EC) were determined using standard procedures (Donagema et al., 2011DONAGEMA, G. K. et al. Manual de métodos de análise de solos. 2ª ed. rev. Rio de Janeiro: Embrapa Solos, 2011. 230p. (Embrapa Solos. Documentos, 132). ).
Samples of soil and aggregates were passed through a 0.210 mm sieve (60-mesh) to determine the TOC by the wet oxidation method with external heating (Yeomans; Bremner, 1988YEOMANS, J. C.; BREMNER, J. M. A rapid and precise method for routine determination of organic carbon in soil. Communications in Soil SciencePlant Analysis, 19(13):1467-1476, 1988. ); the principle of this method is based on the hot oxidation of organic C with potassium dichromate and titration of the remaining dichromate with ammonium ferrous sulfate. To determine the LC content, sub-samples of 1.0 g of soil and aggregate (0.210 mm sieve) were placed in a 50 mL centrifuge tube with 25 mL of KMnO4 solution (0.033 mol L-1). The tubes were agitated at 170 rpm on a horizontal orbital shaker for 1 h and centrifuged at 960 xg for 10 min, always protected from light, with a black bag. After centrifugation, 100 µL (0.1 mL) of supernatant was taken in test tubes, and the volume was made up to 10 mL with deionized water. The LC measurements were made using a spectrophotometer at a wavelength of 565 nm (Shang; Tiessen, 1997SHANG, C.; TIESSEN, H. Organic matter lability in tropical oxisol: Evidence from shifting cultivation, chemical oxidation, particle size, density, and magnetic fractionations. Soil Science , 162(11):795-807, 1997.).
For the determination of humic substances (HS), the soil and aggregates were passed through a 0.210 mm sieve. The extraction was carried out based on the differential solubility of these substances in acids and bases, and they were fractionated based on the methodology of the International Humic Substances Society (IHSS) (Swift, 1996SWIFT, R. S. Method for extraction of IHSS soil fulvic and humic acids. In: SPARKS, D. L. et al. (Eds). Methods of soil analysis: Chemical methods. Madison, Soil Science Society of America, part 3., p.1018-1020, 1996. ). Subsequently, fulvic acid (FA), humic acid (HA), and humin (H) fractions of SOM were obtained. The C content of all fractions was determined based on the wet oxidation method with external heating (Yeomans; Bremner, 1988YEOMANS, J. C.; BREMNER, J. M. A rapid and precise method for routine determination of organic carbon in soil. Communications in Soil SciencePlant Analysis, 19(13):1467-1476, 1988. ).
For X-ray diffractometry, the soil samples and biogenic aggregates were ground and smeared on slides and analyzed using a PANalytical X’Pert Pro PW 3040/60 diffractometer equipped with 1800W tube cobalt (CoKα). The patterns were obtained with a voltage of 40 kV, a current of 40 mA, a step of 0.0008 °2θ s-1, and a scan range from 10 to 60 °2θ (Whiting; Allardice, 1986WHITING, L. D.; ALLARDICE, W. R. X ray diffraction techniques. In: KUTER, A. (Ed.) Methods of soil analysis. Part 1. Physical and mineralogical properties. 2 ed. Madison. American Society of Agronomy, p.331-362, 1986. (Agronomy Series, 9).). The diffraction peaks were identified by utilizing the MINCRYST database (MINCRYST, 2021MINCRYST. Crystallographic and crystallochemical database for minerals and their structural analogues. 2021. Institute of Experimental Mineralogy. Russian Academy of Sciences. WWW-MINCRYST. Available in: <Available in: http://database.iem.ac.ru/mincryst/index.php >. Access in: May, 12, 2021.
http://database.iem.ac.ru/mincryst/index...
).
The data were submitted to analysis of variance by the F test; the results were observed to be significant compared to those obtained by the Tukey test, with a significance of 5%.
RESULTS AND DISCUSSION
Characterization of biogenic aggregates
The biogenic aggregates showed a macroscopic appearance in the form of irregular blocks, with a size greater than 40 mm (Figure 2). Their outer surface was observed to be annular, i.e., convex toward the digestive tract and concave on the opposite side, resulting from the organization formed by juxtaposed segments; its structure was strong to moderate. In the rainy season, when the biogenic aggregates were collected during the daytime, earthworms or any other organisms were not present. However, in nocturnal soil collections, only earthworms were visualized.

Biogenic aggregates of earthworms in the form of irregular blocks. BA1-biogenic aggregates in an area with mesquite tree vegetation (Prosopis juliflora [Sw] D.C); BA2-biogenic aggregates in an area with neem tree vegetation (Azadirachta indica A. Juss); and BA3-biogenic aggregates in an area with forage palm and mesquite tree vegetation (Opuntia sp. and Nopalea sp.).
Regarding the period of formation of biogenic aggregates, it is hypothesized that earthworms, in this semi-arid region, present greater activity at night; this may be due to the milder nighttime climate, specifically the lower relative temperatures and the occurrence of nighttime rains. This fact confirms that earthworm activity is influenced by environmental factors (Brussaard et al., 2012BRUSSAARD, L. et al. Biogeography and phylogenetic community structure of soil invertebrate ecosystem engineers: Global to local patterns, implications for ecosystem functioning and services and global environmental change impacts soil ecology and ecosystem services. In: WALL, D. H. (Ed.). Soil ecology ecosystems services. Oxford University Press, Oxford, p. 201-232, 2012. ), such as temperature (Uvarov; Tiunov; Scheu, 2011UVAROV, A. V.; TIUNOV, A. V.; SCHEU, S. Effects of seasonal and diurnal temperature fluctuations on population dynamics of two Epigeic Earthworm species in forest soil. Soil Biology and Biochemistry , 43(3):559-570, 2011. ; Singh et al., 2020SINGH, J. et al. Climate change effects on earthworms - A review. Soil Organisms, 91(3):113-137, 2020.) and humidity (Kanianska et al., 2016KANIANSKA, R. et al. Assessment of relationships between earthworms and soil abiotic and biotic factors as a tool in sustainable agricultural. Sustainability, 8(9):906, 2016.; Singh et al., 2020SINGH, J. et al. Climate change effects on earthworms - A review. Soil Organisms, 91(3):113-137, 2020.).
The soil classification system, including the Brazilian one, takes into account the activity of organisms in the soil morphological description, but it does not incorporate biological aggregates into the system. The action of pedo-fauna can modify morphogenetic characteristics of soil, such as soil structure, and promote bioturbation (Maaß; Caruso; Rillig, 2015MAAß, S.; CARUSO, T.; RILLIG, M. C. Functional role of microarthropods in soil aggregation. Pedobiologia , 58(2-3):59-63, 2015. ; Gong et al., 2019GONG, X. et al. Earthworms modify soil bacterial and fungal communities through enhancing aggregation and buffering pH. Geoderma , 347:59-69, 2019.; Teshajonova; Valihanov, 2020TESHAJONOVA, M. M.; VALIHANOV, A. O. Some information about earthworms and their biology. Academicia: An International Multidisciplinary Research Journal, 10(12):1171-1176, 2020. ). When excavating the soil, earthworms deposit mucus rich in nutrients and polysaccharides that act as an agglutinating agent. This results in the sedimentation of soil mineral particles, which contributes to the formation of aggregates or macrostructures in the soil (Gong et al., 2019GONG, X. et al. Earthworms modify soil bacterial and fungal communities through enhancing aggregation and buffering pH. Geoderma , 347:59-69, 2019.; Guhra et al., 2020GUHRA, T. et al. Earthworm mucus contributes to the formation of organo-mineral associations in soil. Soil Biology and Biochemistry, 145:107785, 2020. ).
Physicochemical attributes of biogenic aggregates and soil toposequence
The granulometric distribution revealed that, in general, the activity of earthworms resulted in a greater selectivity of particles of smaller diameters. Biogenic aggregates showed a greater tendency to increase the clay fraction with the clay content in BA1, BA2, and BA3 being 17.4%, 15.2%, and 15.5%, respectively, which is approximately similar to the content of P3-A Inceptisol (Table 1). The small increase in the amount of clay in biogenic aggregates is due to soil aggregation and the selection of finer particles as they pass through the earthworm intestine, as reported by Teshajonova and Valihanov (2020TESHAJONOVA, M. M.; VALIHANOV, A. O. Some information about earthworms and their biology. Academicia: An International Multidisciplinary Research Journal, 10(12):1171-1176, 2020. ).
The specific adsorption of earthworm mucus constituents to soil minerals leads to the formation of mucus-mineral associations. These associations contribute to the retention of organic substances from earthworm mucus in soil (micro-) aggregates, which explains the altered physicochemical properties of earthworm-formed aggregates in comparison to the earthworm unaffected “bulk” soil material (Guhra et al., 2020GUHRA, T. et al. Earthworm mucus contributes to the formation of organo-mineral associations in soil. Soil Biology and Biochemistry, 145:107785, 2020. ). Studies show that earthworms can process up to 25% of the Ah horizon in one year; thus, because of the excretion of these biogenic aggregates on the soil surface or subsurface, earthworms constitute important aggregate-forming agents (Lee, 1985LEE, K. E. Earthworms: Their relationships with soils and land use. Academic Press, Australia, 1985. 411p.).
It is important to note that despite the increase in clay content, the biogenic aggregates in this study were made up of sandy soil particles originating from their parent soil materials of syenite-granite. In soils with slow weathering, such as Inceptisol, a greater similarity between soil and soil parent materials has been observed. Thus, the high amounts of sand, mainly coarse sand, favored the low proportion of fine soil particles in the biogenic aggregates. Soil particle size fraction is an important determiner for the retention of SOM, principally for withholding mineral-associated organic carbon (MAOC). Rodrigues Pinto et al. (2021RODRIGUES PINTO, L. A. S. et al. Physicogenic and biogenic aggregates under different management systems in the Cerrado region, Brazil. Revista Brasileira de Ciência do Solo , 45:e0200114, 2021. ) observed that biogenic aggregates have higher MAOC levels than physicogenic aggregates. The results of the present study may be explained by the formation of SOM physical fractions, which are associated with silt and, particularly, clay fractions. The adsorption of organic acids on clay mineral fractions results in chemical protection that enhances the stability of the TOC in the microaggregates (Burak; Fontes; Becquer, 2011BURAK, D. L.; FONTES, M. P. F.; BECQUER, T. Microagregados estáveis e reserva de nutrientes em Latossolo Vermelho sob pastagem em região de cerrado. Pesquisa Agropecuária Tropical, 41(2):229-241, 2011.).
For the chemical attribute, no significant differences in pH were detected between the biogenic aggregates and horizon A soil samples of Inceptisol; either low acidity or weak alkalinity was observed, with pH ranging between 6.82 to 7.28 (Table 2). Soil with low weathering in the semi-arid region usually tends to have close to neutral pH values. Marinho et al. (2016MARINHO, A. C. C. S. et al. Organic matter and physicochemical attributes of a Cambisol under different agricultural uses in a semi-arid region of Brazil. Australian Journal Crop Science, 10(1):32-41, 2016. ), researching Inceptisol in the semi-arid region, reported pH values ranging from 7.1 to 7.9 in soil layers from 0.0 to 0.10 m. However, Mergen Junior et al. (2019aMERGEN JUNIOR, C. A. et al. Atributos químicos em agregados biogênicos e fisiogênicos de solo submetido à aplicação com dejetos suínos. Revista Brasileira de Ciências Agrárias, 14(1):1-8, 2019a.), analyzing biogenic aggregates, reported lower values, with pH ranging between 5.37 to 6.61; these results may be due to the higher levels of organic matter.
For electrical conductivity (EC), there were no differences between the horizon A soils, with values ranging from 0.03 to 0.08 dS m-1; these values are well below the values reported by Marinho et al. (2016MARINHO, A. C. C. S. et al. Organic matter and physicochemical attributes of a Cambisol under different agricultural uses in a semi-arid region of Brazil. Australian Journal Crop Science, 10(1):32-41, 2016. ), who observed a variation from 0.11 to 0.29 dS m-1. However, evaluation of biogenic aggregates revealed higher EC values, i.e., between 0.16 to 0.17 dS m-1 (Table 2). This may be due to the increase in Ca levels verified in biogenic aggregates. Earthworms can alter the environment in which they find themselves, either by the great diversity of organic acids that are released or synthesized in the decomposition process or by the excretion of calcium carbonate (CaCO3) from their calciferous glands, located in the pharynx (Lee, 1985LEE, K. E. Earthworms: Their relationships with soils and land use. Academic Press, Australia, 1985. 411p.; Lambkin et al., 2011LAMBKIN, D. C. et al. Production and dissolution rates of earthworm-secreted calcium carbonate. Pedobiologia, 54:S119-S129, 2011. ). Thus, in addition to affecting the pH, they make the environment more alkaline, and with higher levels of basic cations and salts, they promote changes in the soil of origin. All these factors together may have contributed to the increase in EC in these aggregates (Fiuza; Kusdra; Furtado, 2011FIUZA, S. S.; KUSDRA, J. F.; FURTADO, D. T. Caracterização química e atividade microbiana de coprólitos de Chibui bari (oligochaeta) e do solo adjacente. Revista Brasileira de Ciência do Solo, 35(3):723-728, 2011.).
No Al3+ was detected in the soil or the aggregates. This is possibly associated with pH values above 6.8, causing precipitation of Al3+, and also due to the high content of exchangeable bases.
For nutrients, when analyzing the levels of phosphorus, higher values were observed in the P3-A soil sample (252.4 mg dm-3) compared to the others (Table 2). However, lower phosphorous levels (71.5, 81.0, and 102.2 mg dm-3 for BA1, BA2, and BA3, respectively) were observed in the biogenic aggregates collected from this location, with the highest values observed in the aggregates associated with the vegetal cover of forage palm and mesquite. Possibly this occurred due to the formation of less soluble calcium phosphates in aggregates. For potassium and sodium contents, there was no significant difference among the samples studied (both biogenic aggregates and horizon A samples), with the mean values observed to be 0.71 and 0.81 cmolc dm-3 for K+, and 0.04 and 0.05 cmolc dm-3 for Na+ for the toposequence soil samples and the biogenic aggregates, respectively (Table 2).
Higher levels of Ca2+ were observed in the biogenic aggregates, with BA1, BA2, and BA3 obtaining values of 8.57, 8.14, and 9.89 cmolc dm-3, respectively. There was no difference between the Inceptisol toposequence soil samples, with a mean of 5.2 cmolc dm-3 (Table 2). For Mg2+, within the biogenic aggregates, the highest content was found in BA2 (2.01 cmolc dm-3). In the toposequence soil samples, Mg2+ mobility was observed with enrichment in the P3-A and P4-A parts of the landscape.
The increase in calcium may be a consequence of the excretion of CaCO3 from the calcareous glands in the earthworm pharynx during the ingestion of the soil (Lambkin et al., 2011LAMBKIN, D. C. et al. Production and dissolution rates of earthworm-secreted calcium carbonate. Pedobiologia, 54:S119-S129, 2011. ). Silva Neto et al. (2010SILVA NETO, L. F. et al. Atributos físicos e químicos de agregados pedogênicos e de coprólitos de minhocas em diferentes classes de solos da Paraíba. Ciência e Agrotecnologia, 34(6):1365-1371, 2010. ) reported that the mean levels of Ca2+ and Mg2+ in biogenic aggregates of earthworms were higher than those found in four soil classes (Oxisols, Ultisols, Alfisols, and Nitisols) in Areia city, Paraíba, which is a semi-arid region. Mergen Junior et al. (2019aMERGEN JUNIOR, C. A. et al. Atributos químicos em agregados biogênicos e fisiogênicos de solo submetido à aplicação com dejetos suínos. Revista Brasileira de Ciências Agrárias, 14(1):1-8, 2019a.) found no significant difference in Ca2+ contents when comparing biogenic and physicogenic aggregates with a mean value of 7.07 and 7.25 cmolc kg-1, respectively (0-5 cm), values lower than those found in this study.
For Mg2+, the horizontal and vertical movement of water may have influenced its leaching to the P3-A and P4-A regions; this possibly occurs because Mg2+ has greater mobility in the soil due to its smaller ionic radius. Higher levels of Mg2+ were reported by Mergen Junior et al. (2019aMERGEN JUNIOR, C. A. et al. Atributos químicos em agregados biogênicos e fisiogênicos de solo submetido à aplicação com dejetos suínos. Revista Brasileira de Ciências Agrárias, 14(1):1-8, 2019a.) in biogenic aggregates (5.29 cmolc kg-1) compared to physicogenic aggregates (3.81 cmolc kg-1), which are higher than those found in this study. Normally, the total levels of P, K, Na, Ca, Fe, Cu, Mo, Zn, and Mn, and pH values of biogenic aggregates are higher than the levels observed in soils without the influence of pedo-fauna (Fiuza; Kusdra; Furtado, 2011FIUZA, S. S.; KUSDRA, J. F.; FURTADO, D. T. Caracterização química e atividade microbiana de coprólitos de Chibui bari (oligochaeta) e do solo adjacente. Revista Brasileira de Ciência do Solo, 35(3):723-728, 2011.; Loss et al., 2017LOSS, A. et al. Carbon, nitrogen and natural abundance of 13C and 15N in biogenic and physicogenic aggregates in a soil with 10 years of pig manure application. Soil Tillage and Research, 166:52-58, 2017.). The magnitude of these effects of earthworms on the soil is considerable as they are known to ingest a quantity of soil between 200- and 400-t ha-1 year-1 (Lavelle; Rangel; Kanyonyo, 1983LAVELLE, P.; RANGEL, P.; KANYONYO, J. Mucus production by two species of tropical earthworms: Millsonia lamtoiana (Megascolecidae) and Pontoscolex corethrurus (Glossoscolecidae). In: LEBRUM, P. et al. (Eds.). New trends in soil biology. Louvain-la-Neuve: Dieu-Brichard, p.405-410, 1983. ).
Organic matter fractions of biogenic aggregates and soil toposequence
In general, the biogenic aggregates showed significantly higher mean values for all organic fractions, when compared to horizon A soil samples of Inceptisol toposequence (Table 3). It was observed that the vegetation cover interfered with the contents of TOC in biogenic aggregates, with the highest TOC values found in the aggregates from the area with forage palm and mesquite tree vegetation (BA3-17.8 g kg-1), followed by the area with mesquite tree vegetation (BA1-14.5 g kg-1), and the area with neem tree vegetation (BA2-11.2 g kg-1). In the toposequence soil samples, these values ranged from 5.4 to 7.4 g kg-1 (Table 3).
The biogenic aggregates collected from the areas with mesquite tree and forage palm vegetation (BA3) and only mesquite tree vegetation (BA1) showed the presence of partially decomposed roots, leaves, and thin branches, which may have contributed to the increase in TOC in these aggregates. The higher TOC content in biogenic aggregates, in relation to soil, may be related to the fact that earthworms select materials of organic origin for their food, increasing the concentration of TOC and nutrients in their coprolites (Le Bayon et al., 2017LE BAYON, R. C. G. et al. Earthworms as ecosystem engineers: A review. Earthworms: Types, Roles and Research, NOVA Science Publishers, New York p. 129-178, 2017. ). In addition, the activities of earthworms in the soil contribute to the fragmentation of litter and the transformation of more complex organic materials mixed with mineral particles, complexed with mucus (Vidal et al., 2019VIDAL, A. et al. Earthworm cast formation and development: A shift from plant litter to mineral associated organic matter. Frontiers in Environmental Science, 7:55, 2019.; Guhra et al., 2020GUHRA, T. et al. Earthworm mucus contributes to the formation of organo-mineral associations in soil. Soil Biology and Biochemistry, 145:107785, 2020. ).
The same behavior was observed in biogenic aggregates of Chibui bari (a geophagous tropical earthworm), which exhibited higher levels of nutrients (Ca2+, Mg2+, K+, and P) and TOC; increase in microbial respiration, cation exchange capacity, and pH; and reduction in Al3+ levels (Fiuza; Kusdra; Furtado, 2011FIUZA, S. S.; KUSDRA, J. F.; FURTADO, D. T. Caracterização química e atividade microbiana de coprólitos de Chibui bari (oligochaeta) e do solo adjacente. Revista Brasileira de Ciência do Solo, 35(3):723-728, 2011.). Similar results were demonstrated by Loss et al. (2017LOSS, A. et al. Carbon, nitrogen and natural abundance of 13C and 15N in biogenic and physicogenic aggregates in a soil with 10 years of pig manure application. Soil Tillage and Research, 166:52-58, 2017.) and Mergen Junior et al. (2019aMERGEN JUNIOR, C. A. et al. Atributos químicos em agregados biogênicos e fisiogênicos de solo submetido à aplicação com dejetos suínos. Revista Brasileira de Ciências Agrárias, 14(1):1-8, 2019a.), who reported higher levels of TOC in biogenic aggregates compared to physicogenic ones. However, Rodrigues Pinto et al. (2021RODRIGUES PINTO, L. A. S. et al. Physicogenic and biogenic aggregates under different management systems in the Cerrado region, Brazil. Revista Brasileira de Ciência do Solo , 45:e0200114, 2021. ) did not observe differences in TOC content between biogenic and psychogenic aggregates at areas with no-till and conventional tillage systems and Cerrado vegetation. According to them, the biogenic aggregates changed the SOM dynamics, and more labile SOM fractions were detected in biogenic aggregates (particulate organic carbon and free light fraction carbon). These aggregates were more efficient in reducing SOM decomposition rates and increasing soil carbon content.
For the recalcitrant fraction of organic matter in the soil, the highest levels of carbon in humic substances (C-HS) were observed in biogenic aggregates regardless of vegetation cover, and these values were 19.1, 16.7, and 20.1 g kg-1 for BA1, BA2, and BA3, respectively (Table 3). In the horizon A samples of Inceptisol toposequence, these levels of C-HS ranged from 5.5 to 8.6 g kg-1 (Table 3). The higher levels of all carbon fractions may be because of the stabilization of carbon due to microbial interaction and the release of mucus by earthworms (Angst et al., 2019ANGST, G. et al. Earthworms act as biochemical reactors to convert labile plant compounds into stabilized soil microbial necromass. Communications Biology, 2:441, 2019. ).
For three humic fractions, the highest levels of carbon were for the fraction of humin (C-HUM), followed by humic acid (C-HA), and fulvic acid (C-FA). In humic substances, the predominance of humin is due to its strong interaction with mineral matrix by the formation of highly stable organo-mineral complexes on the soil surface. This results in most of the organic matter being linked to this insoluble fraction.
For C-HUM, the highest levels of C were in biogenic aggregates (11.9, 12.8, and 15.8 g kg-1 for BA1, BA2, and BA3, respectively), while the mean value in the toposequence soils was 5.85 g kg-1 (Table 3). In aggregate BA1 (in an area with mesquite), a reduction in C-HUM and an increase in C-HA were observed (5.1 dag kg-1) (Table 3). For C-FA also, the highest levels were observed for biogenic aggregates, with mean values of 2.1, 1.6, and 1.6 g kg-1 for BA1, BA2, and BA3, respectively. The mean value in the toposequence soils for C-FA was 0.58 g kg-1. The enrichment of carbon in biogenic structures indicates selective feeding of organic material by soil organisms, increasing the carbon protection inside the aggregate. This shows the importance of earthworms in the stabilization of aggregates, which stores carbon in the soil (Pulleman et al., 2005PULLEMAN, M. M. et al. Earthworms and management affect organic matter incorporation and microaggregate formation in agricultural soils. Applied Soil Ecology , 29(5):1-15, 2005.). Besides, earthworms play a major role in the biogeochemical cycling of nutrients in the soil (Jouquet et al., 2009JOUQUET, P. et al. Relevance and limitations of biogenic and physicogenic classification: A comparison of approaches for differentiating the origin of soil aggregates. European Journal of Soil Science, 60(6):1117-1125, 2009.) and positively influence the ecology of humification (Fernandes et al., 2017FERNANDES, J. C. F. et al. Caracterização de agregados biogênicos, intermediários e fisiogênicos em áreas sob domínio da Mata Atlântica Brasileira. Revista Caatinga, 30(1):59-67, 2017. ).
Probably the predominance of carbon in insoluble and soluble recalcitrant fractions of biogenic aggregates occurred due to the chemical stabilization of organic matter. This could be explained by the greater release of Ca2+ from the calcareous glands of earthworms, which associates with the soil by forming metallic bridges with acidic groups. This process results in the formation of calcium humate, which is responsible for the stabilization of organic matter. This demonstrates that not only the supply of organic material but also the nutrients are essential for the humification of the SOM. Majzik and Tombácz (2007MAJZIK, A.; TOMBÁCZ, E. Interaction between humic acid and montmorillonite in the presence of calcium ions II. Colloidal interactions: Charge state, dispersing and/or aggregation of particles in suspension. Organic Geochemistry, 38(8):1330-1340, 2007. ), Briedis et al. (2012BRIEDIS, C. et al. Soil organic matter pools and carbon-protection mechanisms in aggregate classes influenced by surface liming in a no-till system. Geoderma, 170:80-88, 2012. ), and Marinho et al. (2016MARINHO, A. C. C. S. et al. Organic matter and physicochemical attributes of a Cambisol under different agricultural uses in a semi-arid region of Brazil. Australian Journal Crop Science, 10(1):32-41, 2016. ) observed an increase in organic matter in the presence of cations in soil. According to Angst et al. (2019ANGST, G. et al. Earthworms act as biochemical reactors to convert labile plant compounds into stabilized soil microbial necromass. Communications Biology, 2:441, 2019. ), earthworms increase the amount of microbial necromass in stable fractions of micro and macroaggregates. According to Fernandes et al. (2017FERNANDES, J. C. F. et al. Caracterização de agregados biogênicos, intermediários e fisiogênicos em áreas sob domínio da Mata Atlântica Brasileira. Revista Caatinga, 30(1):59-67, 2017. ), even though biogenic aggregates exhibit lesser concentrations of TOC, C-HUM, and C-HA, the levels of these substances are higher than those of intermediate and physicogenic aggregates.
In the humidified fractions of the biogenic aggregates collected from the area with mesquite tree vegetation (BA1), there was a reduction in C-HUM and an increase in C-HA; this may be because of the quality of the source material varies according to vegetation (Singh et al., 2017SINGH, P. et al. Nutrient and enzyme mobilization in earthworm casts: A comparative study with addition of selective amendments in undisturbed and agricultural soils of a mountain ecosystem. International Biodeterioration & Biodegradation, 119:437-447, 2017.). The labile plant compounds are the dominant source of microbial products as they are utilized more efficiently; the microbial products of decomposition (humic acid) are the main precursors of stable SOM. In the vermicomposting process, earthworms ingest and subsequently digest part of the organic waste, thereby fractionating and stimulating the activity of other microorganisms leading to the mineralization of nutrients. This process accelerates the transformation of the waste into humidified material (Dominguez; Perez-Lousada, 2010DOMINGUEZ, J.; PEREZ-LOUSADA, M. Eisenia fetida (savigny, 1826) y Eisenia andrei Bouché, 1972 son dos espécies diferentes de Lombrices de tierra. Acta Zoológica Mexicana , 26(2):321-331, 2010. ), which favors the formation of humic acid.
In the biogenic aggregates, there was also a predominance of labile materials. No significant difference was observed in the LC contents of the biogenic aggregates collected from areas with different vegetation covers, with the values being 2.93, 2.49, and 2.90 g kg-1 for BA1, BA2, and BA3, respectively (Table 3). The mean value of the toposequence soils for LC was 1.31 g kg-1. When earthworms feed on organic matter, they add high amounts of easily degradable organic carbon (mucus) to the ingested material. Angst et al. (2019ANGST, G. et al. Earthworms act as biochemical reactors to convert labile plant compounds into stabilized soil microbial necromass. Communications Biology, 2:441, 2019. ) employed spectroscopic techniques to report a consistent decrease in the relative abundance of lipids and a simultaneous increase in the relative abundance of carbohydrates and proteins in soil fractions affected by earthworms. These substances are derived from the mucus excreted by the earthworm, and because of their lability, they are efficiently utilized by the soil microorganisms. Zhang, Zou and Schaefer (2010ZHANG, M.; ZOU, X.; SCHAEFER, D. A. Alteration of soil labile organic carbon by invasive earthworms (Pontoscolex corethrurus) in tropical rubber plantations. European Journal of Soil Biology , 46(2):74-79, 2010.) observed that the presence of an invasive earthworm (Pontoscolex corethrurus) for 20 months increased the LC content, protected by aggregates, in the topsoil (0-0.05 m) by up to 35%.
It is interesting to question the benefits of the process as a whole. Ecosystem engineers, such as earthworms, play key roles in creating habitats for other organisms and controlling their activities through physical and biochemical processes. The biogenic and organic, and organo-mineral structures produced by earthworms accumulate in the soil space to form three-dimensional mosaics of functional domains, which are inhabited by specific communities of smaller organisms (microfauna and mesofauna) that drive soil processes through specific pathways. Research indicates that LC rich in energy may selectively activate microbial populations and trigger priming effects, resulting in the degradation, synthesis, and sequestration of specific organic substrates (Lavelle et al., 2016LAVELLE, P. et al. Ecosystem engineers in a self-organized soil: A review of concepts and future research questions. Soil Science, 181(3-4):91-109, 2016. ).
Mineralogy of biogenic aggregates and soil toposequence
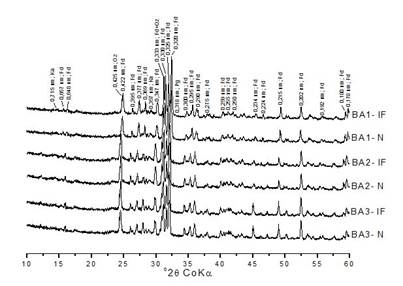
Mineralogical analysis revealed that potassium feldspar (Fd), mostly microcline, was the predominant mineral in the clay fraction of the biogenic aggregates and horizon A soil samples of Inceptisol toposequence (Figure 3, 4, and 5). Apart from this, there was evidence of kaolinite (Ka), with low-intensity peaks at 0.715 and 0.367 nm (Figure 3). Also, quartz (Qz) was identified in P1-A and P2-A areas with a peak of 0.334 nm. Its presence is very common in weathered syenite-granitic rocks, the source material of the studied Inceptisol soils (Figure 4). In general, Ka, plagioclase (Pg), halite (ha), illite (Il), and Qz can be identified in the clay fractions of horizon A samples and biogenic aggregates. The mineral Il was observed at higher concentrations in P3-A and P4-A, with peaks at 1.000 and 0.447 nm. As observed in the case of Ka, Il could have originated by the weathering of K-feldspars (Figure 5). The diffraction peaks characteristic of hematite (0.270; 0.251; and 0.169 nm) and goethite (0.419; 0.244; and 0.269 nm) were absent in the X-ray diffractograms of natural samples. However, the peaks of feldspars and goethite may overlap in some spaces, such as 0.419 nm. The mineral Il was observed more intensely in P3-A and P4-A samples, with peaks at 1,000 and 0,447 nm, and this mineral could have originated by the weathering of the feldspars, similar to kaolinite (Figure 5). In the natural (N) and iron-free (IF) clay fractions, even after separation, fine sand was detected, which may contribute to peaks of primary minerals.

X-ray diffractograms of natural (N) and iron-free (IF) clay fractions of earthworm biogenic aggregates (BA) treated with DCB (dithi onite-citrate-bicarbonate method). Ka-kaolinite and Fd-K-feldspars. BA1-biogenic aggregates in an area with mesquite tree vegetation (Prosopis juliflora [Sw] D.C); BA2-biogenic aggregates in an area with neem tree vegetation (Azadirachta indica A. Juss); and BA3-biogenic aggregates in an area with forage palm and mesquite tree vegetation (Opuntia sp. and Nopalea sp.).

X-ray diffractograms of natural (N) and iron-free (IF) clay fractions of horizon A samples treated with DCB (dithi onite-citrate-bicarbonate method). Il-ilita, Ka-caulinita, Fd-K-feldspars, Qz- quartz, Pg-plagioclásio. P1-A-upper; P2-A-middle third, at a depth of 0-0.20 m.

X-ray diffractograms of natural (N) and iron-free (IF) clay fractions of horizon A samples treated with DCB (dithi onite-citrate-bicarbonate method) Il-ilita, Ka-caulinita; Fd-K- feldspars, Ha-halita (Ha). P3-A-middle third; P4-A-lower slope, at a depth of 0-0.20 m.
The mineralogy of the clay fractions of the biogenic aggregates was observed to be similar irrespective of different vegetation coverings, with a predominance of K-feldspars and traces of kaolinite with low-intensity peaks. The presence of Ka in the samples, even at low-intensity, may be because of the weathering of feldspars (inherited from the parent materials), which probably occurred in more humid climatic conditions than the current ones (Mota et al., 2007MOTA, J. C. A. et al. Atributos mineralógicos de três solos explorados com a cultura do melão na Chapada do Apodi - RN. Revista Brasileira de Ciência do Solo , 31(3):445-454, 2007.; Souza et al., 2010SOUZA, R. V. C. C. et al. Caracterização de solos em uma Topoclimossequência no Maciço de Triunfo - Sertão de Pernambuco. Revista Brasileira de Ciência do Solo , 34(4):1259-1270, 2010. ); however, it was observed that the earthworm activity did not affect the mineral composition of the samples studied. In this specific case, despite being a biogeochemical process, earthworms did not accelerate the mineral weathering (Carpenter et al., 2008CARPENTER, D. et al. The role of earthworm communities in soil mineral weathering: A field experiment. Mineralogical Magazine, 72(1):33-36, 2008. ). Another hypothesis would be some type of organo-mineral protection that has not allowed changes in the mineralogy of the clay fraction. In horizon A Inositol soil samples, the Ka peaks were larger and more intense than in the biogenic aggregates, indicating other pedogenetic processes to be responsible for these factors.
CONCLUSIONS
Results of this study indicate that there is a higher selection of smaller diameter particles in biogenic aggregates compared to horizon A soils of Inceptisol toposequence. The earthworm biogenic aggregates have higher concentrations of Ca+2, TOC, and labile and recalcitrant fractions of organic matter, regardless of the vegetation cover. The action of earthworms did not change the quantity or type of clay minerals. Also, the Ka peaks of horizon A soils of Inceptisol were larger and more intense than those of biogenic aggregates, indicating other pedogenetic processes to be responsible for these factors.
ACKNOWLEDGMENT
We thank sci-edit.com for the English editing of the article.
REFERENCES
- ALVARES, C. A. et al. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22(6):711-728, 2013.
- ANGST, G. et al. Earthworms act as biochemical reactors to convert labile plant compounds into stabilized soil microbial necromass. Communications Biology, 2:441, 2019.
- BITYUTSKII, N. P.; MAIOROV, E. I.; ORLOVA, N. E. The priming effects induced by earthworm mucus on mineralization and humification of plant residues. European Journal of Soil Biology, 50:1-6, 2012.
- BROWN, G. G.; DOMÍNGUEZ, J. Uso das minhocas como bioindicadoras ambientais: princípios e práticas - o 3° Encontro Latino Americano de Ecologia e Taxonomia de Oligoquetas (ELAETAO3). Acta Zoológica Mexicana, 26(2):1-18, 2010.
- BRIEDIS, C. et al. Soil organic matter pools and carbon-protection mechanisms in aggregate classes influenced by surface liming in a no-till system. Geoderma, 170:80-88, 2012.
- BRUSSAARD, L. et al. Biogeography and phylogenetic community structure of soil invertebrate ecosystem engineers: Global to local patterns, implications for ecosystem functioning and services and global environmental change impacts soil ecology and ecosystem services. In: WALL, D. H. (Ed.). Soil ecology ecosystems services. Oxford University Press, Oxford, p. 201-232, 2012.
- BURAK, D. L.; FONTES, M. P. F.; BECQUER, T. Microagregados estáveis e reserva de nutrientes em Latossolo Vermelho sob pastagem em região de cerrado. Pesquisa Agropecuária Tropical, 41(2):229-241, 2011.
- CARPENTER, D. et al. The role of earthworm communities in soil mineral weathering: A field experiment. Mineralogical Magazine, 72(1):33-36, 2008.
- DOMINGUEZ, J.; PEREZ-LOUSADA, M. Eisenia fetida (savigny, 1826) y Eisenia andrei Bouché, 1972 son dos espécies diferentes de Lombrices de tierra. Acta Zoológica Mexicana , 26(2):321-331, 2010.
- DONAGEMA, G. K. et al. Manual de métodos de análise de solos. 2ª ed. rev. Rio de Janeiro: Embrapa Solos, 2011. 230p. (Embrapa Solos. Documentos, 132).
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Sistema Brasileiro de classificação de solos. 5. ed. rev. e ampl. − Brasília, DF: Embrapa, 2018. 356p.
- FERNANDES, J. C. F. et al. Caracterização de agregados biogênicos, intermediários e fisiogênicos em áreas sob domínio da Mata Atlântica Brasileira. Revista Caatinga, 30(1):59-67, 2017.
- FIUZA, S. S.; KUSDRA, J. F.; FURTADO, D. T. Caracterização química e atividade microbiana de coprólitos de Chibui bari (oligochaeta) e do solo adjacente. Revista Brasileira de Ciência do Solo, 35(3):723-728, 2011.
- FUSARO, S. et al. Soil Biological Quality Index based on earthworms (QBS-e). A new way to use earthworms as bioindicators in agroecosystems. Ecological Indicators, 93:1276-1292, 2018.
- GAMA-RODRIGUES, E. F. et al. Atributos físicos, químicos e microbiológicos dos solos sob diferentes usos em topossequências no Noroeste Fluminense. Revista Brasileira de Ciências Agrarias, 13(3):e5543, 2018.
- GONG, X. et al. Earthworms modify soil bacterial and fungal communities through enhancing aggregation and buffering pH. Geoderma , 347:59-69, 2019.
- GUHRA, T. et al. Earthworm mucus contributes to the formation of organo-mineral associations in soil. Soil Biology and Biochemistry, 145:107785, 2020.
- HALLAM, J.; HODSON, M. E. Impact of different earthworm ecotypes on water stable aggregates and soil water holding capacity. Biology and Fertility of Soils, 56:607-617, 2020.
- IUSS WORKING GROUP WRB. 2015. World reference base for soil resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. Rome, Italy: FAO, 2015. 203p.
- JOUQUET, P.; BLANCHART, E.; CAPOWIEZ, Y. Utilization of earthworms and termites for the restoration of ecosystem functioning. Applied Soil Ecology, 73:34-40, 2014.
- JOUQUET, P. et al. Relevance and limitations of biogenic and physicogenic classification: A comparison of approaches for differentiating the origin of soil aggregates. European Journal of Soil Science, 60(6):1117-1125, 2009.
- KANIANSKA, R. et al. Assessment of relationships between earthworms and soil abiotic and biotic factors as a tool in sustainable agricultural. Sustainability, 8(9):906, 2016.
- LAMBKIN, D. C. et al. Production and dissolution rates of earthworm-secreted calcium carbonate. Pedobiologia, 54:S119-S129, 2011.
- LAVELLE, P. et al. Ecosystem engineers in a self-organized soil: A review of concepts and future research questions. Soil Science, 181(3-4):91-109, 2016.
- LAVELLE, P.; RANGEL, P.; KANYONYO, J. Mucus production by two species of tropical earthworms: Millsonia lamtoiana (Megascolecidae) and Pontoscolex corethrurus (Glossoscolecidae). In: LEBRUM, P. et al. (Eds.). New trends in soil biology. Louvain-la-Neuve: Dieu-Brichard, p.405-410, 1983.
- LE BAYON, R. C. G. et al. Earthworms as ecosystem engineers: A review. Earthworms: Types, Roles and Research, NOVA Science Publishers, New York p. 129-178, 2017.
- LEE, K. E. Earthworms: Their relationships with soils and land use. Academic Press, Australia, 1985. 411p.
- LOSS, A. et al. Carbon, nitrogen and natural abundance of 13C and 15N in biogenic and physicogenic aggregates in a soil with 10 years of pig manure application. Soil Tillage and Research, 166:52-58, 2017.
- MAAß, S.; CARUSO, T.; RILLIG, M. C. Functional role of microarthropods in soil aggregation. Pedobiologia , 58(2-3):59-63, 2015.
- MAJZIK, A.; TOMBÁCZ, E. Interaction between humic acid and montmorillonite in the presence of calcium ions II. Colloidal interactions: Charge state, dispersing and/or aggregation of particles in suspension. Organic Geochemistry, 38(8):1330-1340, 2007.
- MARINHO, A. C. C. S. et al. Organic matter and physicochemical attributes of a Cambisol under different agricultural uses in a semi-arid region of Brazil. Australian Journal Crop Science, 10(1):32-41, 2016.
- MEIRELES, H. T. et al. Relações solo-paisagem em topossequência de origem basáltica. Pesquisa Agropecuária Tropical , 42(2):129-136, 2012.
- MELO, M. F. et al. Vermicompostagem: Conversão de resíduos orgânicos em benefícios para solo e plantas. In: ANDRADE, D. F. (Ed.). Tópicos em Ciências Agrárias. Belo Horizonte, Minas Gerais: Poisson, v. 6, p. 35-46, 2020.
- MELO JÚNIOR, H. B. et al. Efeito da ação decompositora da minhoca californiana (Lumbricus rubellus) na composição química de um fertilizante organomineral. Bioscience Journal, 28(1):170-178, 2012.
- MERGEN JUNIOR, C. A. et al. Atributos químicos em agregados biogênicos e fisiogênicos de solo submetido à aplicação com dejetos suínos. Revista Brasileira de Ciências Agrárias, 14(1):1-8, 2019a.
- MERGEN JUNIOR, C. A. et al. Caracterização física de agregados do solo submetido a 10 anos de aplicação de dejetos suínos. Revista Brasileira de Ciências Agrárias , 36(1):79-92, 2019b.
- MINCRYST. Crystallographic and crystallochemical database for minerals and their structural analogues. 2021. Institute of Experimental Mineralogy. Russian Academy of Sciences. WWW-MINCRYST. Available in: <Available in: http://database.iem.ac.ru/mincryst/index.php >. Access in: May, 12, 2021.
» http://database.iem.ac.ru/mincryst/index.php - MOTA, J. C. A. et al. Atributos mineralógicos de três solos explorados com a cultura do melão na Chapada do Apodi - RN. Revista Brasileira de Ciência do Solo , 31(3):445-454, 2007.
- PULLEMAN, M. M. et al. Earthworms and management affect organic matter incorporation and microaggregate formation in agricultural soils. Applied Soil Ecology , 29(5):1-15, 2005.
- RODRIGUES PINTO, L. A. S. et al. Physicogenic and biogenic aggregates under different management systems in the Cerrado region, Brazil. Revista Brasileira de Ciência do Solo , 45:e0200114, 2021.
- ROSSI, J. P. et al. Soil properties inside earthworm patches and gaps in a tropical grassland. European Journal of Soil Biology , 42(1):S284-S288, 2006.
- RUIZ, H. A. Incremento da exatidão da análise granulométrica do solo por meio da coleta da suspensão (silte + argila). Revista Brasileira de Ciência do Solo , 29(5):297-300, 2005.
- SANTOS, R. D. et al. Manual de descrição e coleta de solo no campo. 6.ed. rev. e ampl. Viçosa: Sociedade Brasileira de Ciência do Solo, 2013. 100p.
- SILVA, A. C. et al. Relações entre matéria orgânica do solo e declividade de vertentes em toposseqüência de Latossolos do Sul de Minas Gerais. Revista Brasileira de Ciência do Solo , 31(5):1059-1068, 2007.
- SILVA NETO, L. F. et al. Atributos físicos e químicos de agregados pedogênicos e de coprólitos de minhocas em diferentes classes de solos da Paraíba. Ciência e Agrotecnologia, 34(6):1365-1371, 2010.
- SINGH, J. et al. Climate change effects on earthworms - A review. Soil Organisms, 91(3):113-137, 2020.
- SINGH, P. et al. Nutrient and enzyme mobilization in earthworm casts: A comparative study with addition of selective amendments in undisturbed and agricultural soils of a mountain ecosystem. International Biodeterioration & Biodegradation, 119:437-447, 2017.
- SHANG, C.; TIESSEN, H. Organic matter lability in tropical oxisol: Evidence from shifting cultivation, chemical oxidation, particle size, density, and magnetic fractionations. Soil Science , 162(11):795-807, 1997.
- SOIL TAXONOMY. A Basic system of soil classification for making and interpreting soil surveys. Washington, DC: United States Department of Agriculture (USDA), 1999. 886p.
- SOUZA, R. V. C. C. et al. Caracterização de solos em uma Topoclimossequência no Maciço de Triunfo - Sertão de Pernambuco. Revista Brasileira de Ciência do Solo , 34(4):1259-1270, 2010.
- SWIFT, R. S. Method for extraction of IHSS soil fulvic and humic acids. In: SPARKS, D. L. et al. (Eds). Methods of soil analysis: Chemical methods. Madison, Soil Science Society of America, part 3., p.1018-1020, 1996.
- TESHAJONOVA, M. M.; VALIHANOV, A. O. Some information about earthworms and their biology. Academicia: An International Multidisciplinary Research Journal, 10(12):1171-1176, 2020.
- UVAROV, A. V.; TIUNOV, A. V.; SCHEU, S. Effects of seasonal and diurnal temperature fluctuations on population dynamics of two Epigeic Earthworm species in forest soil. Soil Biology and Biochemistry , 43(3):559-570, 2011.
- VÉLE, A. et al. Local and landscape drivers of ant and carabid beetle communities during spruce forest succession. European Journal of Soil Biology , 47(6):349-356, 2011.
- VIDAL, A. et al. Earthworm cast formation and development: A shift from plant litter to mineral associated organic matter. Frontiers in Environmental Science, 7:55, 2019.
- XIANG, H. et al. In situ earthworm breeding to improve soil aggregation, chemical properties, and enzyme activity in Papayas. Sustainability , 10(4):1193-1206, 2018.
- YEOMANS, J. C.; BREMNER, J. M. A rapid and precise method for routine determination of organic carbon in soil. Communications in Soil SciencePlant Analysis, 19(13):1467-1476, 1988.
- WHITING, L. D.; ALLARDICE, W. R. X ray diffraction techniques. In: KUTER, A. (Ed.) Methods of soil analysis. Part 1. Physical and mineralogical properties. 2 ed. Madison. American Society of Agronomy, p.331-362, 1986. (Agronomy Series, 9).
- ZHANG, M.; ZOU, X.; SCHAEFER, D. A. Alteration of soil labile organic carbon by invasive earthworms (Pontoscolex corethrurus) in tropical rubber plantations. European Journal of Soil Biology , 46(2):74-79, 2010.
Publication Dates
-
Publication in this collection
06 Dec 2021 -
Date of issue
2021
History
-
Received
09 July 2021 -
Accepted
19 Oct 2021