
ABSTRACT:
Eucalypts are the most planted hardwood in the world, with 30 million ha. under cultivation. There are approximately 700 species of eucalypts, but more than 90 % of the plantations are concentrated in nine species of the Subgenus Symphyomyrtus in the Sections Exsertaria, Latoangulatae and Maidenaria. Since 2000, with the advancement of biotechnology, many species have been reclassified and regrouped according to their morphological and physiological characteristics, DNA structure and geographic distribution. This work had as its objective the formulation of a critical analysis which presents the main characteristics that will identify the species of Eucalyptus of these new sections. The species are classified mainly according to geographic distribution, where crosses occur; as evolution in the subgenus Symphyomyrtus is preferably reticulated (with backcrosses), which makes it difficult to identify the rate by molecular and morphological markers. Environmental ecological similarity will define whether the hybrid will become a new species or regress to one parent and become an ecotype. The hybrid will become a new species when it has ecological characteristics different from its parents.
Keywords:
Eucalyptus
; Connexentes; Transversae; Globulares; Viminales
Introduction
Eucalypts make up 10 % of the world´s commercial forests, which translates to 30 million ha (FAO, 2015Food and Agriculture Organization of the United Nations [FAO]. 2015. Global Forest Resources Assessment FAO: Rome. Italy. Available at: http://www.fao.org/3/a-i4808e.pdf. [Accessed Mar 27, 2018]
http://www.fao.org/3/a-i4808e.pdf...
; Payn et al., 2015Payn, T.; Carnus, J.M.; Smith, P.F.; Kimberley, M.; Kollert, W.; Shirong, L.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. 2015. Changes in planted forests and future global implications. Forest Ecology and Management 352: 57-67.). There are about 700 species, however, only nine species occupy more than 90 % of the areas planted with eucalypts, E. camaldulensis and E. tereticornis (Section Exsertaria), E. grandis, E. saligna, E. pellita and E. urophylla (Section Latoangulatae), E. dunnii, E. globulus and E. nitens (Maidenaria Section) (Harwood, 2011Harwood, C. 2011. New introduction doing it right. p. 43-54. In: Walter, J., ed. Developing a eucalypt resource: learning from Australia and elsewhere. Wood Technology Research Centre, ChristChurch, New Zealand.; Ritter, 2014Ritter, M. 2014. Field guide to the cultivated eucalypts (Myrtaceae) and how to identify them. Annual Missouri Botanic Garden 99: 642-687.).
In Brazil, Pryor and Johnson’s (1971)Pryor, L.D.; Johnson, L.A.S. 1971. A Classification of The Eucalypts. Australian National University, Canberra, Australia. classification is still used for Eucalyptus. Brooker (2000)Brooker, M.I.H. 2000. A new classification of the genus Eucalyptus L’Her. (Myrtaceae). Australian Systematic Botany 13: 79–148. published a book that has become the reference in eucalypt classification and is subject to annual adjustments and reclassification to incorporate advances in biotechnology and the evolution of eucalypts itself.
Genetic variation within a species generally has a geographic basis, since the processes of adaptation, gene flow and genetic drift act differently in heterogeneous landscapes and can be strongly influenced by the demographics and spatial distribution of populations (Eckert et al., 2008Eckert, C.G.; Samis, K.E.; Lougheed, C. 2008. Genetic variation across species’ geographical ranges: the central-marginal hypothesis and beyond. Molecular Ecology 17: 1170-1188.). Given this context, hybridization can facilitate gene flow in order to homogenize divergent populations and restrict speciation or, conversely, it can generate unique adaptive phenotypes that lead to species divergence (Rieseberg et al., 2003Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazto, T.; Durphy, J.L.; Schwarzbach, A.E.; Donavan, L.A.; Lexer, C. 2003. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 301: 1211–1216.). Knowledge of a species’ population structure is essential to an understanding of its evolutionary history, as well supporting species improvement and management programs. Therefore, these parameters are important to the classification of a species.
This new classification has three basic pillars, the geographical distribution, where hybridizations can occur; the botanical characteristics of the species, which are considered reflections of the genetic variations between species, and the molecular analyses that generally confirm these genetic variations that are the result of phenotypic variations. We consider natural environments since parental species are adapted according to their evolutionary histories and a very small fraction of hybrids are able to establish themselves due to increased exposure to attack by pests and diseases. In exotic environments, hybrids and pure species compete equally and the success rate of hybridization is much higher.
This work aims to analyze the most important characteristics of these new series and present a practical guide for the differentiation and classification of Eucalyptus species from the main Symphyomyrtus sections.
Materials and Methods
Metadata of species of the subgenus Symphyomyrtus, mainly of the Exsertaria, Latoangulatae and Maidenaria sections, were analyzed taking into account geographic aspects (location of the sections and or series), molecular and botanical markers [plant characteristics, type of bark (Figures 1 to 3), leaves (Figures 4 to 7) (cotyledons, seedling, juvenile, intermediate and adult), flowers (Figures 12 to 17) (position, shape, number of fruits, peduncle, stamens, anthers), ovules, fruits (Figures 8 to 11) (shape, disc and valves), seeds (Figures 18 to 20) (tegument, shape, color, edge, hilum), heartwood, grain and basic density]. These characteristics are a practical guide for the identification of species within the series in each Symphyomyrtus section since it is based on the idea that the majority of species in each series or section, and species that diverged from the majority are considered ecotypes.

Bark texture: 1 = smooth (persistent); 2 = gums decorticated on ribbons; 3 = rough and persistent; 4 = rough deciduous in patches; 5 = tessellated decorticated in small square or rectangular patches.

Bark color: 1 = white; 2 = cream; 3 = yellow; 4 = pink; 5 = red; 6 = brown; 7 = green; 8 = gray; 9 = black; 10 = Orange; 11 = multicolored.

Bark types: 1 = granular; 2 = stringy bark; 3 = ironbark; 4 = bloodwood; 5 = black butt; 6 = ribbon gum; 7 = massmate; 8 = fibrous; 9 = smooth; 10 = peppermint; 11 = box; 12 = compacted; 13 = mattled bark; 14 = tessellated; 15 = rainbow; 16 = scribbes

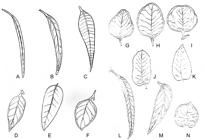
Leaf shape: A = linear; B = narrow lanceolate; C = lanceolate; D = oblong lanceolate; E = wide lanceolate; F = ovoid lanceolate; G = ovoid; H = elliptical; I = orbicular; J = oblong; K = crenated margin; L = falcate; M = oblique; N = cordiform. Source: Adapted from Brooker and Kleinig, 2006.



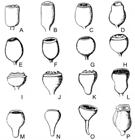
Fruit shape: A = oblong; B = cylindrical; C = barrel; D = Urceolar; E = ovoid; F = truncated ovoid; G = globular; H = truncated globular; I = hemispheric; J, K = widely tapered; L = bell-shaped; M = piriform; N, O = conical; P = quadrangular. Source: Adapted from Brooker and Kleinig, 2006.




Arrangement of inflorescences (2): 1 = in axil of leaves; 2 = in corymbs, in axil of leaves; 3 = in panicles, in axil of leaves; 4 = doubles or decussate in axil of leaves; 5 = individual groups (isolated on the branch); 6 = terminal groups; 7 = isolated flowers in the middle of the branch.
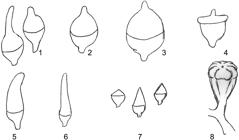
Operculum shape: 1 = rostrate; 2 = hemispherical; 3 = hemispherical apiculate; 4 = flattened; 5 = horn shaped; 6 = elongated; 7= conical; 8 = angular to warty or umbonate.




Seeds shape (1): 1 = saucer or flattened disc; 2 = cuboid or pyramidal; 3 = linear; 4 = in the shape of a boat; 5 = ellipsoidal with wing; 6 = spherical; 7 = ovoid.

Seeds edge (2): 1 = smooth; 2 = cross-linked; 3 = edge sharply toothed; 4 = wrinkled or lacunose; 5 = honeycomb surface.

Results and Discussion
Symphyomyrtus forms a monophyletic group with Telocalyptus and is the largest subgenus of Eucalyptus with about 470 species in 15 Sections, from shrubs to trees over 85 m in height. They occupy a range of zones from arid areas to high open forests with heavy rainfall, and from tropical to temperate climates. They can be found throughout Australia and the islands to the north, Timor, New Guinea, Indonesia and the Philippines. They present the following: seeds with one tegument composed of two layers of cells, considered derivatives, although four cells thick and seeds with two integuments in Angiosperms are more common; the ventral hilum, which will give rise to hemitropic ovules, but there are exceptions; ovules, arranged in four, six, eight or 10 longitudinal lines at the bottom of the placenta which are predominantly hemitropic. All species in this subgenus have two opercules from sepals and petals. The adult leaves are discolored, usually lighter on the abaxial face, with transverse venation and anthers dehiscent by longitudinal slits or sparse pores (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; Drinnan and Ladiges, 1989Drinnan, A.N.; Ladiges, P.Y. 1989. Corolla and androecium development in some Eudesmia eucalypts. (Myrtaceae). Plant Systematic Evolution 65: 239-254.; Ladiges and Humphries, 1983Ladiges, P.Y.; Humphries, C.J. 1983. A cladistic study of Arillastrum, Angophora, and Eucalyptus (Myrtaceae). Botanic Journal of Linnean Society 87: 105-134.; Rozefelds, 1996Rozefelds, A.C. 1996. Eucalyptus phylogeny and history: a brief summary. Tasforests 8: 15-26.).
The Exsertaria Section differs from the other Symphyomyrtus sections in that it presents concolorous intermediate leaves, while in the other sections they are discolorous on the abaxial face, except for the Maidenaria, Platysperma and Incognitae sections, which are also concolorous. Only the Symphyomyrtus section has more than 11 fruits per umbel together with E. robusta of Latoangulatae section. The operculum has the shape of a horn, which is also seen in a number of series of the Latoangulatae and Pumilio section. The exceptions are the Connexentes series and Rostratae subseries. The stamens are erect in all series and subseries. The valves are always exserted, though in the other Symphyomyrtus sections they may not be; the other sections that can also present exserted valves are Platysperma, Latoangulatae, Pumilio and Maidenaria. Seeds with one tegument, except in the Rostratae subseries (E. camaldulensis), which have two integuments, the outer tegument being crystalline, are unique to the subgenus Symphyomyrtus. The seeds are cuboid to pyramidal, with toothed edges, except in the Rostratae Subseries, which is in the form of a flat disc and smooth edge, brown, yellow or black. The heartwood is reddish to brownish.
The Exsertaria section (Table 1) is found throughout continental Australia excluding Tasmania (TAS), in both temperate and tropical regions, in medium to low fertility soils. It presents trees of low or high forest open, and mallees, with a maximum height of 10 m; the bark is quite variable from totally smooth to totally fibrous or rough, deciduous in strips, irregular or regular plates (tessellated, deciduous in square plates or rectangular as in E. brassiana), or persistent bark in different colors with gray, cream, yellow, white and brown. Predominant. All sections have a lignotuber. Cotyledons are bilobed, reniform or oblong, and the leaves show long petioles at all stages. In the Subexsertae subseries, petioles are short in seedlings and juvenile leaves. The leaves are alternated except in the seedlings, that are in opposites, and in the Liberivalvae series this is also found in the juvenile leaves. The shape varies depending on the species, with lanceolate and ovoid shapes predominating. The leaves of the seedlings are discolored on the abaxial surface, except in the Liberivalvae series, which is concolorous in all phases; and in the Subexsertae subseries, which is concolorous (E. bigalerita) or slightly discolored on the abaxial face (E. platyphylla) in seedlings and juvenile leaves and concolorous in the intermediate phase. The adult leaves are concolorous, except in the Connexentes series, which are discolorous in E. major and E. propinqua. The intramarginal veins are sparse to moderate reticulate. The main rib is penniviened, and dense to very densely reticulate; the oil glands are in islands, except for the Subexsertae subseries, which is intersectional. The inflorescence is simple and auxiliary, sprouting from the leaf axilla, with seven to 15 flowers per umbel, with a long peduncle, except in the Subexsertae subseries, which can be sessile or with a short peduncle. The peduncle is flat except in the Liberivalvae series and subseries Exsertae subseries, which are smooth and have a scar on the inflorescence bulb. The operculum is predominantly horn–shaped, but can be rostrate, pointed hemispherical, or conical with continuous ring stamens, erect, which can be flexible or not. The anthers are dorsofixed (flexible), dehiscent by longitudinal slits, oblong or cuboid; the ovules are hemitropous in six vertical rows, except for E. major and E. propinqua where they are in four rows. Long pedicellate or sessile fruits, the shape varies according to the species, with a hemispheric shape predominating. The disc is medium to wide, ascending the hypanthium or fruit, except in the Connexentes series which is level or ascending and in the Subexsertae subseries where it is descending to ascending the hypanthium. Exserted valves (above the disc), in the Subexsertae subseries, are slightly exserted. There are three to five valves per fruit, and a terminal hilum, except in the Connexentes series and Subexsertae subseries, where it can be ventral. The heartwood may be reddish, brownish or pink, and the grain is predominantly interlocked. The basic density in Australia is from 701 to 1100 kg m−3. In commercial forests, cut for five to 20 years, the density decreases by 40 to 60 %.
The Connexentes series (E. hallii, E. broviniensis, E. major and E. propinqua) (small fruited grey gums) are found on the central coast of New South Wales (NSW) up to the Blackdown Tableland in the middle of Queensland (QLD). The bark is smooth, with a granular texture, and deciduous in strips. When new, it has yellow spots, oranges etc. Adult leaves are bright green; discolorous; obscure to dense, with intramarginal parallel veins that are just within the margin. They also present composite inflorescence, inflexible stamen and, seed cuboid to obliquely pyramidal which is brown with a ventral hilum (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Liberivalvae Series (E. bancroftii, E. disclusa, E. interstans, E. prava, E. parramattensis and E. seena) can be found along the middle coast of NSW up to Hervey Bay on the middle coast of QLD. They present the following: trees that reach 15 m in height; bark which is deciduous with granular texture (when it falls orange stains appear); juvenile leaves opposite, petiolate, ovoid to narrow lanceolate; adult leaves alternate, petiolate, opaque, slightly discolorous or concolorous; obscure vein; medium to widely angled; moderate to densely reticulated; inflorescence composed of umbels, with seven to 15 flowers per umbel; ovule in six vertical rows; operculum horn or tapered; erect stamens; anthers cuboid or oblong; versatile, dehiscent by longitudinal slits; valves exerted; seeds cuboid or obliquely pyramidal; toothed edge; brown or black, and terminal hilum (EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
In the Exsertae series (E. brassiana, E. camaldulensis, E. alba, E. rudis, E. amplifolia, E. terrica etc.) there are 40 species and 15 subspecies. They are found in Australia and southern New Guinea. The stem is less colorful and flashy than the species of the Liberivalvae series. They have four subseries: Exsertae, Erythroxylon, Rostratae and Subexsertae. They have a rough, hard and persistent bark and the juvenile leaves are the narrowest in the series (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
).
The Exsertae subseries (Queensland red gums, 8 spp., E. brassiana, E. exserta, E. gillenii, E. nudicaulis etc.) (Phaeoxylon by Brooker, 2000Brooker, M.I.H. 2000. A new classification of the genus Eucalyptus L’Her. (Myrtaceae). Australian Systematic Botany 13: 79–148.) which can be found from northern NSW up to southern New Guinea, including central Australia, occupy the niche of the transition between the mallee (E. morrisii) and the low open forest (E. brassiana). They present the following: smooth, non–powdery, persistent or annually deciduous bark; linear to lanceolate juvenile leaf; adult leaf with moderate to sparse venation; inflorescence sprouting in double or decussate, or single in E. camaldulensis and E. amplifolia; erect stamen; robust fruit; terminal hilum (Hill and Johnson, 2000Hill, K.D.; Johnson, L.A.S. 2000. Systematic studies in the Eucalypts 10: new tropical and subtropical eucalypts from Australia and New Guinea (Eucalyptus - Myrtaceae). Telopea 8: 503-539.; Jones et al., 2016Jones, R.C.; Nicolle, D.; Steane, D.A.; Vaillancourt, R.E.; Potts, B.M. 2016. High density, genome-wide markers and speciose lineage of Eucalyptus. Molecular Phylogenetics and Evolution 105: 63-85.; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Erythroxylon Subseries (Oriental red gums, 14 spp. and 5 subspecies, E. tereticornis) ranges from the Great Eastern Range Division to Flinders Ranges in South Australia (SA) and from the north to the south of New Guinea. It presents smooth, non–powdery, persistent or annually deciduous bark (E. blakelyi, E. glaucina, E. kabiana and E. tereticornis), as well as thin bark (E. chloroclada and E. terrica have thick bark); linear to orbicular juvenile leaf; moderate to sparse tertiary reticulate in adult leaf; inflorescence sprouting in double or decussate; erect stamens; horn–shaped operculum; cuboid to pyramidal seeds, tooted edge, and black; and terminal hilum (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Rostratae subseries (River red gums, 2 spp., 9 subspecies, E. camaldulensis and E. rudis) can be found throughout Australia following the temporary and permanent rivers, and in the floodplains. They prefer soaked soils and present the following: smooth, non–powdery, persistent or annually deciduous bark; juvenile leaves orbicular or lanceolate; adult leaves with moderate to dense venation; inflorescence sprouting in the leaf axils in double (decussed) or single, depending on the subspecies; erect and inflexible stamens in West Australia (WA); rostrate operculum; smooth, yellow disc–shaped seeds, unique in the section; terminal hilum. The seed with two integuments is unique to the section and subgenus Symphyomyrtus (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.). For Jones et al. (2016)Jones, R.C.; Nicolle, D.; Steane, D.A.; Vaillancourt, R.E.; Potts, B.M. 2016. High density, genome-wide markers and speciose lineage of Eucalyptus. Molecular Phylogenetics and Evolution 105: 63-85. E. rudis is contained in E. camaldulensis, a sign that it was an emancipated subspecies.
The Subexsertae subseries (Tropical red gums, 16 spp., 2 subspecies, E. alba, E. bigalerita, E. herbertiana, E. phaltyphylla, E. tintinnas etc.) can be found in the tropical north in Australia and the islands in northern Australia, including New Guinea and Timor. The subseries presents the following: smooth, powdery white bark, annually deciduous; juvenile leaves orbicular or lanceolate; adult leaves with dense to extremely dense venation; oil gland in the form of inter–sectoral star, unique in the Exsertaria series, in the other series they are in the form of islands; inflorescence sprouting in the leaf axils in double (decussed) or single; inflexible stamens, curved or erect; operculum rostrato or pointed hemispherical; cuboid to pyramidal or ovoid seed, toothed edge, and brown; ventral hilum (EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Latoangulatae section (Table 2) (or Transversaria under the informal classification of Pryor and Johnson, 1971Pryor, L.D.; Johnson, L.A.S. 1971. A Classification of The Eucalypts. Australian National University, Canberra, Australia.), can be found in tropical to subtropical climates with high precipitation, in the WA and northeast of Australia, in the mountain ranges of QLD and NSW, and southeast of Gippsland in the territory of Victoria (VIC). They also grow in adjacent coasts, and on the islands to the north, New Guinea, Timor and Indonesia. They are composed of 13 species, from mallees (E. scias) to trees that can exceed 85 m in height (E. grandis), known in Australia as eastern blue gums, red mahoganies and grey gums, characterized by adult leaves discolorous on the underside, which are found only in the sections Pumilio and Inclusae, Connexentes series of the Exsertaria and Deaneanae section, Neglectae and Crenulatae series of the Maidenaria section: Penniviened, which only happens in the Pumilio, Incognitae and Inclusae sections, and in the Connexentes series of the Exsertaria section: secondary and tertiary reticulation of leaves are dense.
The Latoangulatae section presents low or high open forest trees, and mallees, with lignotuber, except E. grandis. The bark can be totally smooth to totally fibrous (E. urophylla) or rough (E. pellita, E. resinifera). The leaves are petiolate; alternate; except in the seedlings, which are opposite for the first to sixth pair; and discolorous on the abaxial face in all phases; the Robustae series does not present intermediate leaves. The leaves are lanceolate, wide lanceolate or ovate in all phases, except the Robustae Series, which can present falcate leaves in the adult phase. Adult leaves are peninerval with dense tertiary reticulate and oil glands in islands. The Latoangulatae section presents the following: simple and auxiliary inflorescence, except in E. grandis, E. urophylla and E. orophila, which can be double; with seven to 15 flowers per umbel; long peduncle markedly flattened leaving a scar on the floral bulb; operculum is predominantly rostrate but can be tapered (E. scias); or in the shape of a horn (E. resinifera); the anthers are erect; predominantly cuboid or oblong (Annulares) or kidney–shaped (Transversae); fixed on the back; versatile, opening through a longitudinal slit; ovules in four to eight vertical rows; bilobed cotyledons; fruits with long pedicel to sessile; the shape varies according to the species; valves leveled to exsertas; seed pyramidal to cuboid, or ovoid; toothed edge; brown or red; reddish heartwood. It is composed of three series: Robustae, Annulares and Transversae (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; Brooker, 2000Brooker, M.I.H. 2000. A new classification of the genus Eucalyptus L’Her. (Myrtaceae). Australian Systematic Botany 13: 79–148.; Brooker and Kleinig, 2006Brooker, M.I.H.; Kleinig, D.A. 2006. Field Guide to Eucalypts. 3ed. Inkata Press, Sydney, Australia.; EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Robustae series species can be found in regions with heavy rain, in northeastern Australia to the middle of the coast in NSW, as well as in the islands to the north, including New Guinea, Indonesia and Timor. They present the following: smooth bark and cream in E. urophylla at altitudes below 800 m; or fibrous only at the base or in the entire stem, reddish brown in E. urophylla at altitudes above 1000 m; leaves with petiole in all stages; leaves of seedlings opposite to sub opposite; alternate in other phases; ovate to broad lanceolate or lanceolate in all phases and in the adult also falcate. They do not have intermediate leaves. Inflorescence is simple and auxiliary, umbels with seven to 15 flowers per umbel, with relatively long peduncle and pedicel, inflexible stamens and relatively large fruit, disk included to excluded; excluded valves; seed pyramidal to cuboid, toothed edge, brown; terminal hilum (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Annulares series species are found on the eastern coast of Australia, from southern NSW up to the east of the Cape York Peninsula in QLD. They present the following: bark is persistent, fibrous, reddish brown; the leaves are broad lanceolate to ovate in all phases except in the seedlings that are lanceolate; flowers with relatively short peduncle and pedicel; horn–shaped, rostrate or hemispheric operculum. E. scias may have originated from the hybridization of one of these species with E. botryoides or E. saligna from the Transversae series (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Transversae series species are found in areas of intense rain on the east coast of Australia, from Gippsland, Victoria, north of Windsor Tableland, north of QLD. Bark totally smooth (E. grandis) to totally fibrous (E. botryoides) or fibrous only at the base (E. grandis, E. saligna), annually descending; relatively small adult leaf, lanceolate, broad lanceolate or ovoid shapes; relatively small fruits, with relatively short peduncles and pedicels; ovules arranged in four vertical rows. Other classifiers placed E. botryoides in the Annulares series, however, its occurrence is quite similar to that of E. saligna, therefore, it was included in this series (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; EUCLID, 2020EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
https://apps.lucidcentral.org/euclid/tex...
; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The Maidenaria section (Table 3) is different from the other Symphyomyrtus sections, in that it presents seedlings with sessile leaves, except in the Bridgesiana and Semiunicolores series, which may have a short petiole; the Foveolatae series, which may have a long petiole, and in the Deaneanae series, the seedling leaves have long petioles in spiral. The Tasmaniae series and subseries Circulares present concolorous juvenile leaves, which only occurs in the Adnataria and Racemus sections in Symphyomyrtus. In the Recurvae series, the adult leaves have no visible venation. It has one fruit per umbel (E. globulus subspecies globulus and E. vernicosa), three fruits per umbel (E. bicostata, E. pseudoglobulus, series Tasmaniae, Semiunicolores and Recurvae, and Circulares subseries) or seven to 15 flowers per umbel, operculum predominantly hemispherical pointed; predominantly ovoid seeds, in the Deaneanae and Semiunicolores series and Euglobulares subseries can be cuboid or pyramidal, wrinkled and brown, or black.
Maidenaria is a large section composed of 13 series, four subseries, 79 species and 29 subspecies which can be found in the areas of heavy precipitation in southeast Australia, especially in the temperate Alps in the Eyre Kangaroo Peninsula in South Australia, up to the west of Brisbane in the south of QLD, and in the less humid areas they form shrubs (mallees). They present the following: lignotubers in all series except Remanentes which is present in the heavy precipitation regions of NSW, VIC and TAS; deciduous bark annually: taxa characterized by bilobed or oblong cotyledons: sessile seedling leaves, except in the Deaneanae, Foveolatae, Crenulatae and Semiunicolores series. The seedling leaves are predominantly ovoid; juvenile leaves ovoid and elliptical; intermediate leaves ovoid and broad lanceolate; and adult leaves lanceolate and narrow lanceolate. The mature crown presents predominantly juvenile and intermediate leaves in the Deaneanae, Neglectae, Crenulatae and Recurvae series. Adult leaves are densely reticulated; oil glands predominantly on islands but may be absent in the Foveolatae series or obscure in the Tasmaniae series. Inflorescences are simple, axillary, except in E. quadrangulata and E. crenulata which are double; anthers erect; dorsofixed; oblong or cuboid; versatile; dehiscent by longitudinal slits; the stamens are inflexible or irregularly inflexible, and all Tasmanian species have inflexible stamens. There is a scar on the floral bulb except in the Sturgissianae series. They also present the following: ovules arranged in four vertical rows fruit sessile to long pedicellate; disc narrow to wide; descending to above; three to four valves per fruit; enclosed to exserted; Ventral hilum, except in the Deaneanae series, which is by inclusion; heartwood predominantly yellow, straight or interlocked grain, with a basic density from 500 to 1100 kg m−3 in native forests and from 250 to 600 kg m−3 in commercial forests (Boland et al., 2006Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.; Brooker, 2000Brooker, M.I.H. 2000. A new classification of the genus Eucalyptus L’Her. (Myrtaceae). Australian Systematic Botany 13: 79–148.).
There is an abundance of hybridization in the genus Eucalyptus, of the same series: the greater the frequency and percentage of success of the hybrids. Symphyomyrtus is the largest subgenus of eucalyptus and the Maidenaria section is one of the largest and most interesting for the study of hybridization, as it occurs in Tasmania, which has mountain ranges where clinal variations thrive with different species, although hybridization is found in all subgenera in Australia and other countries. The average time of differentiation between species is between three and 10 million years, and in sections between 21 and 31 million years, that is, in three million years the crossing capacity decreases approximately 54 %, due to the snowball effect, meaning that they increase negative epistatic interactions leading to incompatibility between species. At least 21 million years are required for the isolation of the sections (Crisp et al., 2011Crisp, M.D.; Burrows, G.E.; Cook, L.G.; Thornhill, A.H.; Bowman, D.M.J.S. 2011. Flammable biomes dominated by eucalypts originated at the Cretaceous-Palaeogene boundary. Nature Communications 2: 193. https://doi.org/10.1038/ncomms1191
https://doi.org/10.1038/ncomms1191...
; Larcombe et al., 2015Larcombe, M.J.; Holland, B.; Steane, D.A.; Jones, R.C.; Nicolle, D.; Vaillancourt, R.E.; Potts, B.M. 2015. Patterns of reproductive isolation in eucalyptus: a phylogenetic perspective. Molecular Biology and Evolution 32: 1833-1846.). During the Quaternary age (17 million years) Tasmania was conjoined to the Australian continent due to the low sea level (120 m below the current level), thus, the species of the continent came into contact with those of the islands and the crossings occurred in practically all temperate species of Symphyomyrtus and Eucalyptus (Monocalyptus), resulting in a predominantly reticulated evolution in Tasmania, as reported by several authors (Jones et al., 2013Jones, R.C.; Steane, D.A.; Lavery, M.; Vaillancourt, R.E.; Potts, B.M. 2013. Several evolutionary processes drive the patterns of genetic differentiation in a complex of forest species. Ecology and Evolution 3: 1-17.; McKinnon et al., 2010McKinnon, G.E.; Smith, J.J.; Potts, B.M. 2010. Recurrent nuclear DNA introgression accompanies chloroplast DNA exchange between two eucalypt species. Molecular Ecology 19: 1367-1380.; Nicolle and Jones, 2018Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.).
The populations are classified according to the area of occurrence as generalists when their range exceeds 600 km; regional, when they occur in a range of 150 to 600 km; and local, when the distribution range is less than 150 km. Generalist populations show a higher average number of alleles by species and population, and polymorphism per locus, heterogozity, and genetic diversity are all higher within populations and lower between populations when compared to regional and local populations, on account of greater gene flow (Kainer et al., 2018Kainer, D.; Stone, E.A.; Padovan, A.; Foley, W.J.; Kulheim, C. 2018. Accuracy of genomic prediction for foliar terpene traits in Eucalyptus polybractea. G3: Genes, Genomes, Genetics 8: 2573-2593.; Jones et al., 2002Jones, R.C.; Steane, D.A.; Potts, B.M.; Vaillancourt, R.E. 2002. Microsatellite and morphological analysis of Eucalyptus globulus populations. Canadian Journal of Forest Research 32: 69-66.; Moran, 1992Moran, G.F. 1992. Patterns of genetic diversity in Australian tree species. New Forests 6: 49-66.; Murray et al., 2019Murray, K.; Janes, J.; Bothwell, H.; Jones, A.; Andrew, R.; Borevit, J. 2019. Landscape drives of genomic diversity and divergence in woodlands Eucalyptus. Molecular Ecology 28: 522-545.; Silva Jr. and Grattapaglia, 2015Silva Júnior, O.B.; Grattapaglia, D. 2015. Genome-wide patterns of recombination, linkage disequilibrium and nucleotide diversity from pooled resequencing and single nucleotide polymorphism genotyping unlock the evolutionary history of Eucalyptus grandis. New Phytologist 208: 830-845.; Supple et al., 2018Supple, M.A.; Jason, G.B.; Broadhurst, A.B.N.; Byrne, M.; Andrew, R.L.; Widdup, A.; Aitken, N.C.; Borevitz, J.O. 2018. Landscape genomic prediction for restoration of a Eucalyptus foundation species under climate change. eLife 7: e31835. https://doi.org/10.7554/eLife.31835.001.
https://doi.org/10.7554/eLife.31835.001...
). When populations are fragmented or isolated, they suffer flooding by pollen from the species most adapted to the site, resulting in hybridization (Field et al., 2008Field, D.L.; Ayre, D.J.; Whelen, R.J.; Young, A.G. 2008. Relative frequency of sympatric species influences rates of interspecific hybridization, seed production and seedling performance in the uncommon Eucalyptus aggregata. Journal of Ecology 96: 1198-1210.; McKinnon et al., 2001McKinnon, G.E.; Vaillancourt, R.E.; Jackson, H.D.; Potts, B.M. 2001. Chloroplast sharing in the Tasmanian eucalypts. Evolution - International Journal of Organic Evolution 55: 703-711.).
Binding imbalance by mutation, hybridization, genetic drift etc. are easier in inbreeding plants and are very rare in generalist populations because the effective population size is very large, favoring crossing between parents, and also because the barriers against inbreeding are stronger than for crossing between different species. Hybridization can facilitate gene flow in order to homogenize divergent populations and restrict speciation or, conversely, it can generate unique adaptive phenotypes that lead to species divergence (Ellstrand and Elam, 1993Ellstrand, N.C.; Elam, D.R. 1993. Population genetic consequences of small population-size: implications for plant conservation. Annual Review of Ecology and Systematics 24: 217–242.; Coyne and Orr, 2004Coyne, J.A.; Orr, H.A. 2004. Speciation. Sinauer, Sunderlande, MA, USA.; Griffin et al., 1988Griffin, A.R.; Burgess, I.P.; Wolf, L. 1988. Patterns of natural and manipulated hybridization in the genus Eucalyptus L’Herit.: a review. Australian Journal of Botany 36: 41-66.; Pollock et al., 2015Pollock, L.J.; Bayly, M.J.; Vesk, P.A. 2015. The roles of ecological and evolutionary processes in plant community assembly: the environment, hybridization and introgression influence co-ocurrence of Eucalyptus. The American Naturalist 185: 784-796.; Rieseberg et al., 2003Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazto, T.; Durphy, J.L.; Schwarzbach, A.E.; Donavan, L.A.; Lexer, C. 2003. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 301: 1211–1216.; Shepherd and Raymond, 2010Shepherd, M.; Raymond, C. 2010. Species differentiation and gene flow in Blackbutts (genus Eucalyptus Subgenus Section of Eucalyptus Pseudophloius). Conservation Genetics 11: 1965–1978.; Steane et al., 2006Steane, D.A.; Conod, N.; Jones, R.C.; Vaillancourt, R.E.; Potts, B.M. 2006. A comparative analysis of population structure of a forest tree, Eucalyptus globulus (Myrtaceae), using microsatellite markers and quantitative traits. Tree Genetics and Genomes 2: 30-38.). The size of the isolated population and the environmental conditions determined which path was followed. The speciation process is gradual and continuous, and there is no point of divergence in speciation, which makes it difficult to identify taxa because there are homologous characters (similarity due to common ancestry), and homoplasia (similarity due to convergence). However, speciation or differentiation in relation to parents increases with geographical distance. This past and present reticulated evolution makes it difficult to identify by molecular and morphological markers, and, thus, there is a fine line between ecotypes and new species (Bradburd et al., 2018Bradburd, G.S.; Coop, G.M.; Ralph, P.L. 2018. Inferring continuous and discrete genetic structure in space. Genetics 210: 33-52.; Rutherford et al., 2018Rutherford, S.; Rossetto, M.; Bragg, J.G.; McPherson, H.; Benson, D.; Bonser, S.P.; Wilson, P.G. 2018. Speciation in the presence of gene flow: population genomics of closely related and diverging Eucalyptus species. Heredity 121: 126-141.).
Another very important factor is exclusion by reproductive affinity. Two species from the same series, which theoretically would cross more easily and, therefore, make the co–occurrence of this hybrid more common, rarely occurs or the occurrence is less than expected. This is because the closer the species are genetically, the greater the competition between both and the greater the susceptibility to pests and diseases, therefore the hybrid between these species has less chance of surviving, since susceptibility to pests and diseases would be the sum of the attacks suffered by the parents, which would reduce their chances of survival. In genetically more distant species, for example, from different sections, the set of pests and diseases is different, and the defense mechanisms are also different, so that there is morphological similarity, such as specific leaf are, for example, there is hybridization and backcrossing, and the hybrids share these defense mechanisms. The competition between pure and hybrid species is less than if the species were genetically close. Nearby species only occurred in the same area if there were reproductive isolation, in, for example, different flowering seasons.
In exotic environments, such as Brazil, genetically close species can cross more easily, due to the proximity of the species, and there are not so many pests and diseases as in the natural environments in which they evolved. Thus, the occurrence of natural hybrids between close species is much greater than in the origin of the species.
Conclusions
Hybridization occurs in all Eucalyptus Subgenera. More fragmented populations tend to experience greater flooding of pollen because this is one of the best ways to increase their variability. If the species adapts to the site, it will undergo several backcrosses, fixing adaptive genes and becoming an ecotype. If the species does not adapt to the location, the hybrid diverges from the parents to become a new species. There are several examples in the two lines in the past and today, and therefore, we suggest that the hybrid be recognized as a new species when it has morphological and genetic characteristics different from those of the parents. If the parental species show ecological similarity, the same height as the plant or specific leaf area, there will be backcrosses and haplotype sharing, with a tendency for a new ecotype to appear. If the species do not show many ecological similarities, the tendency is for the hybrid to diverge from its parents to become a new species.
References
- Boland, D.J.; Brooker, M.I.H.; Chippendale, G.M.; Hall, N.; Hyland, B.P.M.; Johnston, R.D.; Kleinig, D.A.; McDonald, M.W.; Turner, J.D. 2006. Forest Trees of Australia. 5ed. CSIRO, Collingwood, Australia.
- Bradburd, G.S.; Coop, G.M.; Ralph, P.L. 2018. Inferring continuous and discrete genetic structure in space. Genetics 210: 33-52.
- Brooker, M.I.H. 2000. A new classification of the genus Eucalyptus L’Her. (Myrtaceae). Australian Systematic Botany 13: 79–148.
- Brooker, M.I.H.; Kleinig, D.A. 2006. Field Guide to Eucalypts. 3ed. Inkata Press, Sydney, Australia.
- Coyne, J.A.; Orr, H.A. 2004. Speciation. Sinauer, Sunderlande, MA, USA.
- Crisp, M.D.; Burrows, G.E.; Cook, L.G.; Thornhill, A.H.; Bowman, D.M.J.S. 2011. Flammable biomes dominated by eucalypts originated at the Cretaceous-Palaeogene boundary. Nature Communications 2: 193. https://doi.org/10.1038/ncomms1191
» https://doi.org/10.1038/ncomms1191 - Drinnan, A.N.; Ladiges, P.Y. 1989. Corolla and androecium development in some Eudesmia eucalypts. (Myrtaceae). Plant Systematic Evolution 65: 239-254.
- Eckert, C.G.; Samis, K.E.; Lougheed, C. 2008. Genetic variation across species’ geographical ranges: the central-marginal hypothesis and beyond. Molecular Ecology 17: 1170-1188.
- Ellstrand, N.C.; Elam, D.R. 1993. Population genetic consequences of small population-size: implications for plant conservation. Annual Review of Ecology and Systematics 24: 217–242.
- EUCLID Eucalyptcs of Australia. 2020. Eucalypts of Australia. 4ed. Available at: https://apps.lucidcentral.org/euclid/text/intro/index.html [Accessed Apr 19, 2020]
» https://apps.lucidcentral.org/euclid/text/intro/index.html - Food and Agriculture Organization of the United Nations [FAO]. 2015. Global Forest Resources Assessment FAO: Rome. Italy. Available at: http://www.fao.org/3/a-i4808e.pdf [Accessed Mar 27, 2018]
» http://www.fao.org/3/a-i4808e.pdf - Field, D.L.; Ayre, D.J.; Whelen, R.J.; Young, A.G. 2008. Relative frequency of sympatric species influences rates of interspecific hybridization, seed production and seedling performance in the uncommon Eucalyptus aggregata. Journal of Ecology 96: 1198-1210.
- Griffin, A.R.; Burgess, I.P.; Wolf, L. 1988. Patterns of natural and manipulated hybridization in the genus Eucalyptus L’Herit.: a review. Australian Journal of Botany 36: 41-66.
- Harwood, C. 2011. New introduction doing it right. p. 43-54. In: Walter, J., ed. Developing a eucalypt resource: learning from Australia and elsewhere. Wood Technology Research Centre, ChristChurch, New Zealand.
- Hill, K.D.; Johnson, L.A.S. 2000. Systematic studies in the Eucalypts 10: new tropical and subtropical eucalypts from Australia and New Guinea (Eucalyptus - Myrtaceae). Telopea 8: 503-539.
- Jones, R.C.; Nicolle, D.; Steane, D.A.; Vaillancourt, R.E.; Potts, B.M. 2016. High density, genome-wide markers and speciose lineage of Eucalyptus. Molecular Phylogenetics and Evolution 105: 63-85.
- Jones, R.C.; Steane, D.A.; Lavery, M.; Vaillancourt, R.E.; Potts, B.M. 2013. Several evolutionary processes drive the patterns of genetic differentiation in a complex of forest species. Ecology and Evolution 3: 1-17.
- Jones, R.C.; Steane, D.A.; Potts, B.M.; Vaillancourt, R.E. 2002. Microsatellite and morphological analysis of Eucalyptus globulus populations. Canadian Journal of Forest Research 32: 69-66.
- Kainer, D.; Stone, E.A.; Padovan, A.; Foley, W.J.; Kulheim, C. 2018. Accuracy of genomic prediction for foliar terpene traits in Eucalyptus polybractea. G3: Genes, Genomes, Genetics 8: 2573-2593.
- Ladiges, P.Y.; Humphries, C.J. 1983. A cladistic study of Arillastrum, Angophora, and Eucalyptus (Myrtaceae). Botanic Journal of Linnean Society 87: 105-134.
- Larcombe, M.J.; Holland, B.; Steane, D.A.; Jones, R.C.; Nicolle, D.; Vaillancourt, R.E.; Potts, B.M. 2015. Patterns of reproductive isolation in eucalyptus: a phylogenetic perspective. Molecular Biology and Evolution 32: 1833-1846.
- McKinnon, G.E.; Smith, J.J.; Potts, B.M. 2010. Recurrent nuclear DNA introgression accompanies chloroplast DNA exchange between two eucalypt species. Molecular Ecology 19: 1367-1380.
- McKinnon, G.E.; Vaillancourt, R.E.; Jackson, H.D.; Potts, B.M. 2001. Chloroplast sharing in the Tasmanian eucalypts. Evolution - International Journal of Organic Evolution 55: 703-711.
- Moran, G.F. 1992. Patterns of genetic diversity in Australian tree species. New Forests 6: 49-66.
- Murray, K.; Janes, J.; Bothwell, H.; Jones, A.; Andrew, R.; Borevit, J. 2019. Landscape drives of genomic diversity and divergence in woodlands Eucalyptus. Molecular Ecology 28: 522-545.
- Nicolle, D.; Jones, R.C. 2018. A revised classification for the predominantly eastern Australian Eucalyptus subgenus Symphyomyrtus sections Maidenaria, Exsertaria, Latoangulatae and related smaller sections (Myrtaceae). Telopea 21: 129-145.
- Payn, T.; Carnus, J.M.; Smith, P.F.; Kimberley, M.; Kollert, W.; Shirong, L.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. 2015. Changes in planted forests and future global implications. Forest Ecology and Management 352: 57-67.
- Pollock, L.J.; Bayly, M.J.; Vesk, P.A. 2015. The roles of ecological and evolutionary processes in plant community assembly: the environment, hybridization and introgression influence co-ocurrence of Eucalyptus. The American Naturalist 185: 784-796.
- Pryor, L.D.; Johnson, L.A.S. 1971. A Classification of The Eucalypts. Australian National University, Canberra, Australia.
- Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazto, T.; Durphy, J.L.; Schwarzbach, A.E.; Donavan, L.A.; Lexer, C. 2003. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 301: 1211–1216.
- Ritter, M. 2014. Field guide to the cultivated eucalypts (Myrtaceae) and how to identify them. Annual Missouri Botanic Garden 99: 642-687.
- Rozefelds, A.C. 1996. Eucalyptus phylogeny and history: a brief summary. Tasforests 8: 15-26.
- Rutherford, S.; Rossetto, M.; Bragg, J.G.; McPherson, H.; Benson, D.; Bonser, S.P.; Wilson, P.G. 2018. Speciation in the presence of gene flow: population genomics of closely related and diverging Eucalyptus species. Heredity 121: 126-141.
- Shepherd, M.; Raymond, C. 2010. Species differentiation and gene flow in Blackbutts (genus Eucalyptus Subgenus Section of Eucalyptus Pseudophloius). Conservation Genetics 11: 1965–1978.
- Silva Júnior, O.B.; Grattapaglia, D. 2015. Genome-wide patterns of recombination, linkage disequilibrium and nucleotide diversity from pooled resequencing and single nucleotide polymorphism genotyping unlock the evolutionary history of Eucalyptus grandis. New Phytologist 208: 830-845.
- Steane, D.A.; Conod, N.; Jones, R.C.; Vaillancourt, R.E.; Potts, B.M. 2006. A comparative analysis of population structure of a forest tree, Eucalyptus globulus (Myrtaceae), using microsatellite markers and quantitative traits. Tree Genetics and Genomes 2: 30-38.
- Supple, M.A.; Jason, G.B.; Broadhurst, A.B.N.; Byrne, M.; Andrew, R.L.; Widdup, A.; Aitken, N.C.; Borevitz, J.O. 2018. Landscape genomic prediction for restoration of a Eucalyptus foundation species under climate change. eLife 7: e31835. https://doi.org/10.7554/eLife.31835.001
» https://doi.org/10.7554/eLife.31835.001
Edited by
Publication Dates
-
Publication in this collection
02 Apr 2021 -
Date of issue
2021
History
-
Received
24 June 2020 -
Accepted
09 Oct 2020




















