Resumos
O estudo in vitro foi realizado a partir de sementes de S. allagophylla, colhidas de plantas crescidas na Reserva Biológica de Moji-Guaçu. Foram testadas duas interfaces da adaptação desta espécie às condições do cerrado: efeito do pH e das concentrações de nutrientes, utilizando o meio básico de Murashige & Skoog (MS) e o de Gamborg et al. (B5). Modificações do meio MS foram feitas em relação ao pH, com um gradiente de valores iniciais, indo do 4,2 ao 5,8 (intervalos de 0,2), e em relação aos nutrientes KNO3, KH2PO4 e MgSO4.7H2O, com concentrações progressivamente menores destes. Quanto ao meio B5 foi testada a composição nutricional nas concentrações totais e reduzidas à metade (B5 50%). Os resultados mostraram que as adaptações desta espécie do cerrado in vitro foram: todos os explantes, independente do valor inicial do pH, acidificaram o meio e o crescimento foi mais favorável em meios com menores valores iniciais de pH; o crescimento não foi afetado pela diminuição da concentração de nitrato e a redução da composição nutricional do meio B5 até promoveu o crescimento, principalmente quanto à expansão foliar; o crescimento foi similar tanto na presença como na ausência total de KH2PO4 e de MgSO4.7H2O (em relação ao meio MS). Estes resultados são consistentes com o conceito de uma planta bem adaptada em absorver nutrientes de solos de cerrado, solos estes ácidos, pobres em nutrientes e ricos em alumínio.
In vitro studies on S. allagophylla were carried out using, as a starting material, seeds collected from wild plants growing in "campo cerrado" vegetation at the Reserva Biológica e Estação Experimental de Moji-Guaçu, SP, Brazil. Two aspects of plant adaptation to cerrado conditions were tested in vitro: effect of pH and nutrient concentration, using the two traditional Murashige & Skoog (MS) e o de Gamborg et al. (B5) media. MS basic medium was modified in relation to pH by a gradient of initial pH values ranging from 4.2 to 5.8 at 0.2 pH unit intervals and with progressively lower concentrations of KNO3, KH2PO4 and MgSO4.7H2O. B5 medium was tested with standard or half-strength nutrients. Results shown that, in vitro, this material showed: increased growth in acid media and all explants cultivated in vitro regardless of the initial pH acidified the media; growth was not affected by the reduction of nitrate concentration and even B5 half-strength nutrients promoted growth, mainly in relation to leaf expansion; growth was the same in the complete presence or absence (in relation to the MS medium) of KH2PO4 and MgSO4.7H2O. These in vitro results are consistent with a plant well-adapted to absorbing nutrients in acid, nutrient-poor, aluminium-rich cerrado soils.
In vitro culture; acid pH; cerrado; Gesneriaceae
Estudo de nutrição mineral in vitro relacionado à adaptação de Sinningia allagophylla (Martius) Wiehler (Gesneriaceae) às condições de cerrado
MÁRCIA A. NOVAES GOMES1 1. Departamento de Fisiologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil. e SIMONE L. KIRSZENZAFT SHEPHERD1,2 1. Departamento de Fisiologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970 Campinas, SP, Brasil.
(recebido em 6 de novembro de 1998; aceito em 5 de janeiro de 2000)
ABSTRACT - (In vitro study of mineral nutrition in Sinningia allagophylla (Martius) Wiehler (Gesneriaceae) related to adaptation to cerrado conditions). In vitro studies on S. allagophylla were carried out using, as a starting material, seeds collected from wild plants growing in campo cerrado vegetation at the Reserva Biológica e Estação Experimental de Moji-Guaçu, SP, Brazil. Two aspects of plant adaptation to cerrado conditions were tested in vitro: effect of pH and nutrient concentration, using the two traditional Murashige & Skoog (MS) e o de Gamborg et al. (B5) media. MS basic medium was modified in relation to pH by a gradient of initial pH values ranging from 4.2 to 5.8 at 0.2 pH unit intervals and with progressively lower concentrations of KNO3, KH2PO4 and MgSO4.7H2O. B5 medium was tested with standard or half-strength nutrients. Results shown that, in vitro, this material showed: increased growth in acid media and all explants cultivated in vitro regardless of the initial pH acidified the media; growth was not affected by the reduction of nitrate concentration and even B5 half-strength nutrients promoted growth, mainly in relation to leaf expansion; growth was the same in the complete presence or absence (in relation to the MS medium) of KH2PO4 and MgSO4.7H2O. These in vitro results are consistent with a plant well-adapted to absorbing nutrients in acid, nutrient-poor, aluminium-rich cerrado soils.
RESUMO - (Estudo de nutrição mineral in vitro relacionado à adaptação de Sinningia allagophylla (Martius) Wiehler (Gesneriaceae) às condições de cerrado). O estudo in vitro foi realizado a partir de sementes de S. allagophylla, colhidas de plantas crescidas na Reserva Biológica de Moji-Guaçu. Foram testadas duas interfaces da adaptação desta espécie às condições do cerrado: efeito do pH e das concentrações de nutrientes, utilizando o meio básico de Murashige & Skoog (MS) e o de Gamborg et al. (B5). Modificações do meio MS foram feitas em relação ao pH, com um gradiente de valores iniciais, indo do 4,2 ao 5,8 (intervalos de 0,2), e em relação aos nutrientes KNO3, KH2PO4 e MgSO4.7H2O, com concentrações progressivamente menores destes. Quanto ao meio B5 foi testada a composição nutricional nas concentrações totais e reduzidas à metade (B5 50%). Os resultados mostraram que as adaptações desta espécie do cerrado in vitro foram: todos os explantes, independente do valor inicial do pH, acidificaram o meio e o crescimento foi mais favorável em meios com menores valores iniciais de pH; o crescimento não foi afetado pela diminuição da concentração de nitrato e a redução da composição nutricional do meio B5 até promoveu o crescimento, principalmente quanto à expansão foliar; o crescimento foi similar tanto na presença como na ausência total de KH2PO4 e de MgSO4.7H2O (em relação ao meio MS). Estes resultados são consistentes com o conceito de uma planta bem adaptada em absorver nutrientes de solos de cerrado, solos estes ácidos, pobres em nutrientes e ricos em alumínio.
Key words - In vitro culture, acid pH, cerrado, Gesneriaceae
Introdução
O cerrado constitui-se na segunda maior formação vegetal brasileira, depois da Floresta Amazônica, abrangendo aproximadamente 25% do território brasileiro (Ratter & Ribeiro 1996). A qualidade do solo do cerrado é um fator que determina o tipo de vegetação característico deste ecossistema. O solo é profundo, bem drenado e submetido a intenso intemperismo e lixiviação, o que o torna ácido, pobre em nutrientes e com altos teores de ferro e alumínio. A toxidade do íon alumínio pode ser um dos fatores mais importante que limita o crescimento das plantas em tais solos (Goodland & Ferri 1979). Na planta, o alumínio causa a inibição do seu crescimento, provavelmente por impedir o influxo de minerais como cálcio, magnésio e fosfato, ou por inibir o crescimento da raiz (Meharg 1994).
Estudos realizados por Haridasan (1982) com espécies nativas do cerrado, identificaram algumas delas como acumuladoras de alumínio (acumulando mais de 1000 mg.kg-1). Tal acúmulo, observado nas folhas, não está associado ao baixo nível dos minerais essenciais, indicando que as espécies acumuladoras de alumínio possuem mecanismos de adaptação eficientes na utilização dos nutrientes (Haridasan et al. 1987).
Apesar da alta acidez e baixa fertilidade dos solos, o cerrado está ameaçado por constituir a principal fronteira agropecuária do Brasil, em razão de sua proximidade dos centros consumidores, pela topografia plana e pela estrutura viária satisfatória (Ker et al. 1992). A ocupação do cerrado pelo setor agropecuário, somada ao acentuado extrativismo, concorre para o processo de extinção de espécies, sendo a conservação de fragmentos deste ecossistema de grande importância para a manutenção da ampla biodiversidade de que é formado, a qual não é completamente conhecida. Vários trabalhos revelam o potencial econômico de espécies do cerrado nas áreas de produção de alimentos e industrial, de produção de fármacos e do paisagismo (Uexküll & Mutert 1995). Estudos básicos de ecofisiologia, bioquímica e de biotecnologia, com a micropropagação de plantas nativas, são importantes para um melhor entendimento dos mecanismos de adaptação das espécies a essa formação vegetal.
Sinningia allagophylla (sinonímia Rechsteineria spicata), espécie da família Gesneriaceae, é uma planta herbácea nativa, de ampla distribuição nos cerrados, apresentando caracteres que são considerados comuns aos de plantas adaptadas às condições do cerrado, com o caule ereto, simples, piloso, nascendo de um a vários ramos de um órgão subterrâneo; as folhas resistentes, com inervação proeminente e as flores reunidas em cachos terminais com a corola cor-de-abóbora (Ferri 1969). Estudos fenológicos realizados por Almeida (1994) mostraram que a espécie S. allagophylla é uma planta perene com ciclo anual, que cresce, floresce e, em seguida, tem um gradual declínio, resultando na perda de sua parte aérea durante as estações secas (outono e inverno). As gemas dormentes permanecem nos órgãos subterrâneos, com o brotamento ocorrendo quando as condições se tornam favoráveis (na primavera). Septímio (1994) cita que esta espécie é conhecida como cravo do campo e que suas folhas e flores têm uso popular como depurativo, febrífugo e diurético.
A espécie citada respondeu favoravelmente às condições de cultivo in vitro (Almeida & Shepherd 1999), obtendo-se material, independente da época do ano, para a realização de estudos ecofisiológicos. O objetivo do presente trabalho foi estudar o comportamento in vitro de S. allagophylla frente aos fatores nutricionais, tais como valores do pH e da composição do meio, visando ao embasamento do conhecimento dos mecanismos de adaptação e o aperfeiçoamento do protocolo de micropropagação desta espécie do cerrado.
Material e métodos
O material botânico utilizado nos estudos (sementes e folhas) foi coletado de plantas crescidas em área de campo cerrado, setor destinado à pesquisa não perturbatória, da Reserva Biológica e Estação Experimental de Moji-Guaçu (22°15-16'S e 47°8-12'W, 585-635 m de altitude) no estado de São Paulo. As plantas, a partir das quais foram feitas as coletas, formavam uma população de oito indivíduos distribuídos em duas áreas, cada uma de 5-10 m2, separadas por uma distância de 20-30 m. Amostra local do solo foi coletada a 20 cm abaixo da superfície.
A primeira abordagem do estudo da adaptação de S. allagophylla foi feita in vitro com a observação do efeito do pH do meio e uma segunda abordagem com o estudo nutricional. Para uma visão ecofisiológica mais ampla da adaptação, foram realizadas determinações de nutrientes presentes no material crescido in situ, tanto nas folhas de plantas crescidas no cerrado, como na amostra do solo do local.
Os estudos in vitro foram realizados com plântulas obtidas assepticamente a partir de sementes. As plântulas foram cultivadas em meio de cultura básico de Murashige & Skoog (1962) (MS), com pH ajustado para 5,8. Essa cultura foi mantida durante dois meses em tubos de ensaio de fundo chato com 8 cm de altura e 2,5 cm de diâmetro, submetidos à luminosidade constante de 35,49 mmol.m-2.s-1, fornecida por duas lâmpadas fluorescentes, e a uma temperatura de 25 ± 2 °C. A partir das plântulas, obteve-se segmentos nodais com 0,8 cm de comprimento, com um par de folhas, os quais foram utilizados para os estudos do efeito de diferentes pHs do meio e de diferentes concentrações de nutrientes. Os parâmetros analisados foram: comprimento do eixo principal, número e comprimento das raízes, número de folhas e grau de expansão foliar.
Meio com diferentes valores iniciais de pH - Explantes foram cultivados assepticamente durante 90 dias em meio básico MS em estado líquido, sobre suporte de papel de filtro Whatman no. 1, num gradiente de valores iniciais do pH, os quais foram ajustados antes da autoclavagem, variando de 4,2 a 5,8, em intervalos de 0,2, totalizando nove tratamentos, com cinco repetições cada. A observação da mudança do pH foi possível graças a presença do indicador verde bromocresol (5 mL.L-1) adicionado ao meio quando do seu preparo. Foram preparados controles com água destilada e o indicador, para os diferentes pH, a cada coleta de dados.
Meio com diferentes concentrações de KH2PO4 e MgSO4.7H2O - Explantes foram cultivados assepticamente durante 90 dias em meio MS em estado líquido, utilizando-se como suporte papel de filtro Whatman no. 1, em pH 4,4 e com diferentes combinações de concentrações de KH2PO4 (170, 17, 1,7 e 0,0 mg.L-1) e de MgSO4.7H2O (370, 37, 3,7, 0,0 mg.L-1), totalizando 16 tratamentos com cinco repetições de cada um.
Meio com diferentes concentrações de nutrientes, particularmente para o íon NO3- - Explantes foram inoculados em meios em estado semi-sólido com pH 4,8, totalizando cinco tratamentos com cinco repetições cada um: meio MS (tratamento 1), MS com de KNO3 (tratamento 2), MS com de KNO3 (tratamento 3), meio Gamborg et al. (1968) (B5) (tratamento 4) e B5 com as concentrações nutricionais reduzidas à metade (B5 50%) (tratamento 5). O pH 4,8 foi escolhido por ser mais ácido que o pH 5,8 recomendado para o meio de cultivo e por permitir a geleificação do meio.
Para resultados obtidos referentes aos parâmetros observados foram calculados a média e o desvio padrão e foi realizada análise de variância ANOVA, entre os tratamentos e dentro dos tratamentos.
Para a análise química da planta, foram utilizadas as folhas por serem elas os órgãos que melhor refletem o estado nutricional da planta (Malavolta et al. 1997). Os métodos utilizados exigem uma quantidade mínima de 100 g de matéria seca para ser efetiva, o que inviabiliza a análise do material vegetal cultivado in vitro. As análises foram realizadas no Instituto Agronômico de Campinas, Seção de Fertilidade do Solo e Nutrição de Plantas.
Análise das folhas - Após a lavagem, secagem e moagem das folhas verdes, os extratos para a determinação dos teores dos elementos foram obtidos através da digestão com ácido perclórico, com extração separada para o N, por digestão sulfúrica, e para o Al, por digestão seca (incineração) (Bataglia et al. 1983). Para a determinação dos teores dos elementos foi utilizada a espectrometria de emissão atômica com plasma de argônio (ICP-AES).
Análise do solo - Foram utilizados extratores, como a resina para o P, K, Ca e Mg; água quente para o B (Abreu et al. 1994), solução complexante DTPA para o Zn, Fe, Cu e Mn (Lindsay & Norvell 1978) e o extrator KCl para o Al. A determinação dos teores foi dada por métodos espectrométricos e o valor do pH foi determinado em solução CaCl2 0,1 M.
Resultados
Os diferentes pHs iniciais do meio MS sofreram alterações durante os 90 dias de cultivo, com uma tendência à diminuição após 15 dias da inoculação dos explantes. Logo após a autoclavagem, a diminuição dos valores do pH não foi significativa, pois não ocorreu alteração da coloração dos meios, comparada com o controle. Pela tabela 1 pode-se observar que as diferenças entre os valores iniciais de pH e após o cultivo, foram significativas nos tratamentos com valores iniciais acima de 4,6. Para os tratamentos com pH inicial 4,2 e 4,4 não ocorreram alterações significativas. Os diferentes valores do pH não apresentaram diferenças significativas nos parâmetros analisados, entretanto foi observada uma tendência de maior crescimento dos explantes cultivados no meio com pH inicial 4,6 (dados não mostrados).
Valores iniciais do pH do meio MS e após 90 dias (média ± desvio padrão) do cultivo in vitro de S. allagophylla. Diferenças significativas em nível de 5% (*), 1% (**) e 0,1% (***) (teste de Duncan).
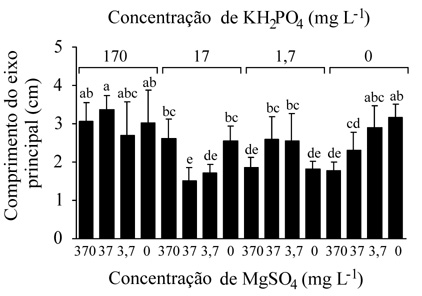
Os valores de pH do meio MS com diferentes concentrações de KH2PO4 e de MgSO4.7H2O, durante os 90 dias do cultivo, não apresentaram variações significativas, mantendo-se entre 4,4 e 4,6. Os resultados do efeito das diferentes combinações de concentrações de KH2PO4 e de MgSO4.7H2O, sobre os parâmetros analisados, evidenciaram maior crescimento dos explantes tanto em meio MS com 100% da concentração de KH2PO4 e de MnSO4.7H2O, quanto em meio MS com a ausência destes nutrientes. Valores significativamente menores, com diferenças em nível de 0,1% (teste de Duncan), para o comprimento do eixo principal (figura 1), para o comprimento das raízes (figura 2) e o número de folhas (figura 3), foram observados nos tratamentos com a combinação de menores concentrações de KH2PO4 e/ou de MnSO4.7H2O e/ou a ausência de apenas um deles.
Efeito de diferentes combinações de concentrações de KH2PO4 e de MgSO4.7H2O em mg.L-1 sobre o comprimento do eixo principal de S. allagophylla, após 90 dias de cultivo (médias e respectivos desvios padrões). Letras diferentes indicam diferenças significativas em nível de 0,1% (teste de Duncan).
Efeito de diferentes combinações de concentrações de KH2PO4 e de MgSO4.7H2O em mg.L-1 sobre o número (A) e o comprimento das raízes (B) de S. allagophylla, após 90 dias de cultivo (médias e respectivos desvios padrões). Classes dos valores dos parâmetros: número de raízes 1 = 0; 2 = 1-5; 3 = 5-10; 4 = 10-15; 5 = acima de 15 raízes e comprimento das raízes 1 = 0,0; 2 = 0,1-1,0 cm; 3 = 1,0-2,0 cm; 4 = 2,0-3,0 cm; 5 = 3,0-4,0 cm. Letras diferentes indicam diferenças significativas em nível de 5% (teste de Duncan) para o número de raízes e de 0,1% para o comprimento.
Efeito de diferentes combinações de concentrações de KH2PO4 e de MgSO4.7H2O em mg.L-1 sobre o número de folhas de S. allagophylla, após 90 dias de cultivo (médias e respectivos desvios padrões). Classes dos valores do parâmetro número de folhas 1 = 0; 2 = 1-5; 3 = 5-10; 4 = 10-15; 5 = acima de 15 folhas. Letras diferentes indicam diferenças significativas em nível de 0,1% (teste de Duncan).
Quanto aos meios com concentrações diferentes de nutrientes, particularmente em relação ao nitrato, no meio B5 50% os explantes apresentaram maior crescimento, com uma tendência para valores mais altos do comprimento do eixo principal, do número e comprimento das raízes e do número de folhas, e com valores significativamente maiores para a expansão foliar (figura 4). A diminuição da concentração de nitrato do meio MS não afetou o crescimento.
Efeito das diferentes concentrações de nutrientes, particularmente do NO3-, sobre a expansão foliar de S. allagophylla, após 90 dias de cultivo (médias e respectivos desvios padrões). Classes dos valores do parâmetro e expansão foliar 1 = pouca expansão (área de 9-38 mm2); 2 = média ex--94 mm2) . Letras diferentes indicam diferenças significativas em nível de 1% (teste de Duncan).
O resultado da análise química da amostra do solo, da Reserva Biológica de Moji-Guaçu, indicou ser um solo extremamente ácido, com o valor do pH em 3,9, com baixos teores de Ca, Mg, Mn e Zn, teores muito baixos para P e K, médios para B e Cu e extremamente altos para o Fe (30,0 mg.dm-3) e alto para o Al trocável (7,08 mmolc.dm-3) (tabela 2). O nível de matéria orgânica apresentou-se dentro de valores considerados médios para solo do cerrado, a capacidade de troca catiônica (CTC) mostrou um valor baixo e a de saturação de bases com um valor extremamente baixo. O resultado da análise química das folhas maduras, coletadas de plantas crescidas nesta reserva, foi de baixo teor de P, teores médios de N e Zn, altos de K, Ca, Mg, B, Cu, Mn e extremamente altos para o Fe (1211,0 mg.kg-1) e para o Al (2991,9 mg.kg-1) (tabela 3). Os resultados foram comparados com a tabela padrão, da composição nutricional considerada adequada, elaborada por Epstein (1975).
Resultado da análise química da amostra do solo de campo cerrado, coletada na Reserva Biológica de Moji-Guaçu. MO = matéria orgânica, H+Al = acidez potencial, SB = soma de bases, CTC = capacidade de troca catiônica, V = saturação de bases.
Resultado da análise química das folhas maduras coletadas de plantas de S. allagophylla, crescidas em campo cerrado na Reserva Biológica de Moji-Guaçu.
Discussão
A diminuição observada no pH do meio de cultivo, após 90 dias, pode estar relacionada à absorção diferencial de NH4+ e NO3- pelos explantes ou a produtos metabólicos excretados para o meio (Caldas et al. 1990). A tendência em diminuir o pH do meio e de maior crescimento em pHs mais baixos, indicou que a espécie estudada tem um pH ótimo para o cultivo in vitro menor que o normalmente recomendado (pH 5,8), sugerindo que a mesma apresenta estratégias de adaptação para crescimento em meios mais ácidos. Sabe-se que baixos valores do pH afetam o crescimento das plantas pelo efeito direto do excesso de hidrogênio e indireto pela mudança na solubilidade de vários elementos minerais importantes (Islam et al. 1980).
O maior crescimento observado nos explantes cultivados em meio MS com 100% da concentração de KH2PO4 e de MgSO4.7H2O e em meio MS com ausência destes, com o maior comprimento do eixo principal e das raízes, do número de raízes e folhas e da expansão foliar, indicou que a espécie S. allagophylla apresenta eficiência na utilização de tais sais presentes nas concentrações do meio MS e que a espécie utiliza os nutrientes armazenados em seus tecidos quando da ausência deles no meio, sugerindo estratégias de economia e utilização de nutrientes. Tais características vêm sendo observadas em estudos com diferentes espécies, onde, sob controle genético, espécies adaptadas aos solos ácidos e pobres, possuem eficiência em utilizar os nutrientes (Baligar et al. 1997). Já os tratamentos com combinações de menores concentrações de KH2PO4 e/ou de MgSO4.7H2O e/ou ausência de um deles resultaram em um crescimento significativamente menor para todos os parâmetros analisados. Talvez as combinações de concentrações de tais sais nestes tratamentos tenham interferido na absorção ou na translocação de outros nutrientes essenciais ao crescimento dos explantes. Várias interações entre os íons estão envolvidas com a absorção e utilização deles pela planta, como a competição, o sinergismo e a relação cátion-ânion (Marschner 1997), sendo importante que os sais se encontrem nos substratos em proporções quantitativas equilibradas (Ziegler 1993). É necessário considerar, na preparação de soluções nutritivas artificiais, não somente os íons requeridos, mas também a razão individual entre eles, ou seja, o balanço iônico (Mohr & Schopfer 1995).
Os resultados obtidos do cultivo em meios com diferentes concentrações de nutrientes, particularmente do nitrato, para todos os parâmetros analisados, não mostraram diferenças significativas quanto à concentração utilizada, o que indica que a espécie tem eficiência para utilizar o nitrogênio e demais nutrientes em todos os tratamentos. O meio B5 50% provavelmente é mais favorável para o seu cultivo in vitro, pois neste meio foi observado um maior crescimento dos explantes, com uma diferença significativamente maior para a expansão foliar.
A análise do solo mostrou que a espécie S. allagophylla cresce em solo extremamente ácido, pobre em nutrientes e bases trocáveis, baixa CTC e com teores de Fe e de Al trocável considerados tóxicos, características essas encontradas em solo de cerrado, devido a sua origem geológica e a ação do intemperismo e lixiviação, que causam a perda por dissolução de Ca2+, Mg2+, K+, PO4 e N, resultando em um aumento da acidez do solo (Goodland & Ferri 1979). Com esta acidez ocorre a remoção dos cátions trocáveis dos locais de troca catiônica e o aumento da solubilidade do íons Al+3 (Malavolta & Kliemann 1985). Os valores absolutos de Al+3 trocável não foram altos (sendo considerados altos acima de 10,0 mmolc.dm-3), entretanto como os valores de Ca2+, Mg2+ e K+ foram muito baixos, os valores relativos do Al+3 trocável são considerados altos e tóxicos. O teor de Al também refletiu que a extrema acidez deste solo não teve como único determinante o íon alumínio, mas provavelmente também o íon H+. A baixa saturação de bases e a CTC não muito reduzida, indicam que provavelmente a matéria orgânica, com teor considerado médio, seja a principal fração envolvida na formação de cargas negativas e troca de cátions neste solo. Resultados semelhantes foram observados em estudos realizados por Lopes (1984), com a análise de diferentes tipos de solos de cerrados.
Pela análise química das folhas verificou-se que os teores dos nutrientes tenderam a valores altos, com exceção do P que apresentou concentração de 0,9 g.kg-1 (adequada é 2 g.kg-1), evidenciando estratégias para alta eficiência de absorção, transporte e utilização da reduzida quantidade dos nutrientes disponíveis. Já os teores extremamente altos do Al e do Fe encontrados nas folhas, evidenciaram que a espécie tolera bem tais elementos, podendo ser considerada uma acumuladora de Al, pois apresentou em suas folhas uma concentração de 2991,9 mg.kg-1. S. allagophylla provavelmente apresenta estratégias de tolerância ao íon alumínio, observadas em outras espécies tolerantes, como a formação de complexos deste íon com ácidos orgânicos, principalmente com citrato e malato (Kochian 1995). Além disso, os altos teores dos nutrientes essenciais presentes em suas folhas demonstraram que essa espécie provavelmente apresenta como forma de tolerância, a capacidade de manter o fluxo de íons e o potencial de membrana nas células das raízes na presença deste íon tóxico (Miyasaka et al. 1989).
Os resultados dos tratamentos in vitro, demonstrando que S. allagophylla tem crescimento favorável em meios ácidos e com baixa concentração de nutrientes, concordam com os resultados obtidos nas análises químicas das folhas e do solo onde crescem plantas dessa espécie. Este trabalho sugere que S. allagophylla possui estratégias de eficiência na obtenção e utilização dos nutrientes em solo pobre e ácido do cerrado.
Agradecimentos - Os autores agradecem ao auxílio técnico da Sra. Dulce Regina Garcia Joaquim, do Departamento de Fisiologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas. M.A.N. Gomes agradece à CAPES e ao CNPq pela Bolsa de Mestrado concedida.
2. Autor para correspondência: kirszenz@obelix.unicamp.br
- ABREU, C.A., ABREU, M.F., RAIJ, B., BATAGLIA, O.C. & ANDRADE, J.C. 1994. Extraction of boron from soil by microwane heating for ICP-AES determination. Communications in Soil Science and Plant Analysis 25:3321-3333.
- ALMEIDA, V.P. 1994. Tuberizaçăo em Sinningia allagophylla (Martius) Wiehler (Gesneriaceae), uma espécie do cerrado. Tese de Doutorado, Universidade Estadual de Campinas, Campinas.
- ALMEIDA, V.P. & SHEPHERD, S.L.K. 1999. Sinningia allagophylla (Gesneriaceae): cultivation in vitro of a native plant of Brazilian cerrado. Revista Brasileira de Botânica 22:381-384.
- BALIGAR, V.C., PITTA, G.V.E., GAMA, E.E.G., SCHAFFERT, R.E., BAHIA, A.F.C.F. & CLARK, R.B. 1997. Soil acidity effects on nutrient use efficiency in exotic maize genotypes. Plant Soil 192:9-13.
- BATAGLIA, O.C., FURLANI, A.M.C., TEIXEIRA, J.P.F., FURLANI, P.R. & GALO, J.R. 1983. Métodos de análise química de plantas (Boletim técnico). Instituto Agronômico de Campinas, Campinas.
- CALDAS, L.S., HARIDASAN, P. & FERREIRA, M.E., 1990. Meios nutritivos. In Técnicas e aplicaçőes da cultura de plantas (A.C. Torres & L.S. Caldas, eds.). ABCTP/EMBRAPA-CNPH, Brasília, p.37-70.
- EPSTEIN, L. 1975. Os componentes inorgânicos das plantas. In Nutriçăo mineral das plantas: princípios e perspectivas. 2 ed. Livros Técnicos e Científicos Editora S.A., Rio de Janeiro, p.50-52.
- FERRI, M.G. 1969. Plantas do Brasil - espécies do cerrado. E. Blucher & EDUSP, Săo Paulo.
- GAMBORG, O.L., MILLER, R.A. & OJIMA, K. 1968. Nutrient requirements of suspension cultures of soybean root cells. Experimental Cell Research 50:151-158.
- GOODLAND, R. & FERRI, M.G. 1979. Relaçőes planta-solo. In Ecologia do cerrado (R. Goodland & M.G. Ferri, eds.). EDUSP, Săo Paulo, p.146-157.
- HARIDASAN, M. 1982. Aluminium accumulation by some cerrado native species of central Brazil. Plant Soil 65:265-273.
- HARIDASAN, M., HILL, P.G. & RUSSELL, D. 1987. Semiquantitative estimates of Al and other cations in the leaf tissues of some Al-accumulating species using electron probe microanalysis. Plant Soil 104:99-102.
- ISLAM, A.K.M.S., EDWARDS, D.G. & ASHER, C.J. 1980. pH optima for crop growth. Plant Soil 54:339-357.
- KER, J.C., PEREIRA, N.R., CARVALHO-JUNIOR, W. & CARVALHO-FILHO, A. 1992. Cerrados: solos, aptidăo e potencialidade agrícola. In Anais do Simpósio sobre manejo e conservaçăo do solo no cerrado, Campinas, p.1-28.
- KOCHIAN, L.V. 1995. Cellular mechanisms of aluminum toxicity and resistance in plants. Annual Review of Plant Physiology and Plant Molecular Biology 46:237-260.
- LINDSAY, W.L. & NORVELL, W.A. 1978. Development of a DTPA soil test for zinc, iron, manganese and copper. Soil Science Society America Journal 42:421-428.
- LOPES, A.S. 1984. Características e propriedades químicas da camada superficial (0-20) dos solos sob Cerrado. In Solos sob Cerrado - Características, propriedades e manejo. Associaçăo Brasileira para Pesquisa da Potassa e do Fosfato, Piracicaba, p.10-60.
- MALAVOLTA, E. & KLIEMANN, H.J. 1985. Deficięncias e excessos. In Desordens nutricionais no Cerrado. Associaçăo Brasileira para Pesquisa da Potassa e do Fosfato, Piracicaba, p.05-65.
- MALAVOLTA, E., VITTI, G.C. & OLIVEIRA, S.A. 1997. Princípios, métodos e técnicas de avaliaçăo. In Avaliaçăo do estado nutricional das plantas. Associaçăo Brasileira para Pesquisa da Potassa e do Fosfato, Piracicaba, p.55-114.
- MARSCHNER, H. 1997. Ion uptake mechanisms of individual cells and roots: short-distance transport. In Mineral nutrition of higher plants. 2 ed. Academic Press, London, p.6-78.
- MEHARG, A.A. 1994. Integrated tolerance mechanisms: constitutive and adaptive plant responses to elevated metal concentrations in the environment. Plant Cell and Environment 17:989-993.
- MIYASAKA, S.C., KOCHIAN, L.V., SHAFF, J.E. & FOY, C.D. 1989. Mechanisms of aluminum tolerance in wheat. Plant Physiology 91:1188-1196.
- MOHR, H. & SCHOPFER, P. 1995. Metabolism of water and inorganic ions. In Plant physiology. Springer-Verlag, Berlin, p.259-267.
- MURASHIGE, T. & SKOOG, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiologia Plantarum 15:473-497.
- RATTER, J.A. & RIBEIRO, J.F. 1996. Biodiversity of the flora of the cerrado. In Anais do VIII Simpósio Sobre o Cerrado, Brasília, p.2-5.
- SEPTÍMIO, L.R. 1994. As plantas: nome vulgar, caracterizaçăo botânica, uso terapęutico. In A fitoterapia brasileira baseada em ervas medicinais do Cerrado. SIPE - Secretaria de Intercâmbio e Projetos Especiais, Brasília, p.27-90.
- UEXKÜLL, H.R. & MUTERT, E. 1995. Global extent, development and economic impact of acid soils. Plant Soil 171:1-15.
- ZIEGLER, H. 1993. Los nutrientes y su transformación en la planta. In Stransburger, tratado de botánica (D.V. Denfer, F. Ehrendorfer, A. Bresinsky & H. Ziegler, eds.). 32 ed. Ediçőes Omega, Barcelona, p.314-370.
Datas de Publicação
-
Publicação nesta coleção
02 Fev 2001 -
Data do Fascículo
Jun 2000
Histórico
-
Recebido
06 Nov 1998 -
Aceito
05 Jan 2000