Resumos
O objetivo deste trabalho foi determinar as concentrações de amido e açúcares solúveis em tecidos de gemas e base de gemas de plantas de pessegueiro (Prunus persica (L.) Batsch) cultivar Jubileu, com e sem sintomas da Morte-Precoce-do-Pessegueiro, durante o período de repouso. Os ramos foram coletados em quatro épocas durante a dormência (11-06, 11-07, 29-07 e 05-08), no inverno de 2003. Foram conduzidos dois experimentos separadamente, um para cada pomar. A elevada concentração de amido nos tecidos das plantas sem sintomas de morte-precoce na fase da saída de dormência propiciou um adequado suprimento energético para que a brotação e a floração destas plantas ocorressem de maneira uniforme e regular. Por outro lado, a antecipação na quebra de dormência das plantas com sintomas da morte-precoce, provocada pelo desencadeamento da síndrome, intensificou a degradação do amido e do sorbitol em ambos os tecidos na saída de dormência, possivelmente, para o fornecimento de glicose e frutose.
Prunus sp; amido; açúcares solúveis; morte-precoce-do-pessegueiro
The aim of this study was to quantify the carbohydrates content (starch and soluble sugars) in bud tissues and bud base of peach trees (Prunus persica (L.) Batsch) cultivar Jubileu in plants with and without Peach-tree-short-life (PTSL) symptoms during the dormant period. The branches were collected in four different times during the dormant period (June, 11th, July, 11th, July, 29th, August, 05th) in the winter of 2003. Two experiments were conducted separately, one for each orchard. The higher concentration of starch in plnt tissues without PTSL symptoms in the final of the dormant period provided a suitable energetic supply , leading to a regular and uniform budbreak of the plants. On the other hand, the budbreak anticipation of the plants with PTSL symptoms, which was provoked by the syndrome starting, intensified the starch and sorbitol breakdown in both tissues in the final of the dormant period, possibly producing glucose and fructose.
Prunus sp.; starch; soluble sugars; Peach-tree-shot-life
DEFESA FITOSSANITÁRIA
Concentrações de carboidratos em tecidos de pessegueiro (Prunus persica (L.) Batsch) cv. jubileu em plantas com ou sem sintomas de morte-precoce durante o período de dormência1 1 (Trabalho 096-2006).
Carbohydrates content in peach trees tissues (Prunus persica (L.) Batsch) cv. jubileu in plants with and without peach-tree-short-life symptoms during the dormant period
Anderson Carlos MarafonI; Flávio Gilberto HerterII; Marcos Antônio BacarinIII; Alexandre Couto RodriguesIV; Valtair VeríssimoV
IDoutorando do Curso de Pós-Graduação em Fisiologia Vegetal, Universidade Federal de Pelotas, Instituto de Biologia, Departamento de Botânica, Cx. Postal 354, 96010-900, Pelotas-RS Correio eletrônico: anderson_marafon@ufpel.edu.br
IIPesquisador da Embrapa Clima Temperado, Br 392, Km 78, Cx. Postal 403, 96001-970, Pelotas-RS. Correio eletrônico: herter@cpact.embrapa.br
IIIDocente do Curso de Pós-Graduação em Fisiologia Vegetal da Universidade Federal de Pelotas, Instituto de Biologia, Departamento de Botânica , Cx. Postal 354, 96010-900, Pelotas-RS Correio eletrônico: bacarin@ufpel.edu.br
IVPesquisador visitante Embrapa Clima Temperado, Br 392, Km 78, Cx. Postal 403, 96001-970, Pelotas-RS. Correio eletrônico: rcale@cpact.embrapa.br
VDoutorando do Curso de Pós-Graduação em Fruticultura, Universidade Federal de Pelotas, Faculdade de Agronomia Eliseu Maciel, Cx. Postal 354, 96001-015 Pelotas-RS Correio eletrônico: valtair_verissimo@yahoo.com.br
RESUMO
O objetivo deste trabalho foi determinar as concentrações de amido e açúcares solúveis em tecidos de gemas e base de gemas de plantas de pessegueiro (Prunus persica (L.) Batsch) cultivar Jubileu, com e sem sintomas da Morte-Precoce-do-Pessegueiro, durante o período de repouso. Os ramos foram coletados em quatro épocas durante a dormência (11-06, 11-07, 29-07 e 05-08), no inverno de 2003. Foram conduzidos dois experimentos separadamente, um para cada pomar. A elevada concentração de amido nos tecidos das plantas sem sintomas de morte-precoce na fase da saída de dormência propiciou um adequado suprimento energético para que a brotação e a floração destas plantas ocorressem de maneira uniforme e regular. Por outro lado, a antecipação na quebra de dormência das plantas com sintomas da morte-precoce, provocada pelo desencadeamento da síndrome, intensificou a degradação do amido e do sorbitol em ambos os tecidos na saída de dormência, possivelmente, para o fornecimento de glicose e frutose.
Termos para indexação: Prunus sp, amido, açúcares solúveis, morte-precoce-do-pessegueiro.
ABSTRACT
The aim of this study was to quantify the carbohydrates content (starch and soluble sugars) in bud tissues and bud base of peach trees (Prunus persica (L.) Batsch) cultivar Jubileu in plants with and without Peach-tree-short-life (PTSL) symptoms during the dormant period. The branches were collected in four different times during the dormant period (June, 11th, July, 11th, July, 29th, August, 05th) in the winter of 2003. Two experiments were conducted separately, one for each orchard. The higher concentration of starch in plnt tissues without PTSL symptoms in the final of the dormant period provided a suitable energetic supply , leading to a regular and uniform budbreak of the plants. On the other hand, the budbreak anticipation of the plants with PTSL symptoms, which was provoked by the syndrome starting, intensified the starch and sorbitol breakdown in both tissues in the final of the dormant period, possibly producing glucose and fructose.
Index Terms: Prunus sp., starch, soluble sugars, Peach-tree-shot-life.
INTRODUÇÃO
Nos últimos anos, vem se intensificando a ocorrência de um tipo de depauperamento de pessegueiros no Rio Grande do Sul designado Morte-Precoce-do-Pessegueiro. Trata-se de uma síndrome que se caracteriza por um colapso da planta durante a floração, ou no início da brotação. A brotação e a floração das plantas são anormais, podendo ocorrer brotações tardias no interior da copa e nos ramos mais vigorosos (Fortes & Osório, 2003).
A síndrome é conseqüência de uma série de fatores desencadeantes, que predispõem ou enfraquecem a planta, associados às oscilações bruscas da temperatura no período de quebra da dormência. Dentre os fatores predisponentes, temos: deficiência nutricional ou toxidez em razão do excesso de nutrientes; solos ácidos (pH < 6,0); solos encharcados ou com deficiência hídrica; ferimentos nas raízes decorrentes de práticas culturais; podas prematuras, e uso de porta-enxerto suscetível ao nematóide-anelado (Mesocriconema xenoplax [Raski] Loof & DeGrisse) (Medeiros & Raseira, 1998).
Campos & Carvalho (1994) verificaram que, sob condições de estresse hídrico, os teores de carboidratos solúveis totais são reduzidos drasticamente após a retirada dos frutos em plantas com sintomas de morte-precoce. Devido ao estresse, as plantas em dormência, debilitadas após um período de baixa atividade fotossintética, manifestam sintomas de deficiência nutricional, morte de ramos e paralisação do crescimento das gemas e, em condições de oscilações bruscas de temperatura, aquelas mais debilitadas morrem.
O objetivo desta pesquisa foi determinar as concentrações de amido e açúcares solúveis em tecidos de gemas e base de gemas de pessegueiro (Prunus persica (L.) Batsch) cultivar Jubileu, em plantas com e sem sintomas da síndrome da Morte-Precoce-do-Pessegueiro, durante o período de dormência.
MATERIAIS E MÉTODOS
Foram conduzidos dois experimentos separadamente: Experimento 1 - plantas sem sintomas e Experimento 2 - plantas com sintomas da Morte-Precoce-do-Pessegueiro.
Cada experimento foi conduzido em pomares separados, ambos com três anos de implantação, situados acerca de 5Km de proximidade entre si, no município de Pelotas-RS. O clima da região de Pelotas, conforme a classificação de Köppen, é do tipo Cfa: subtropical com precipitação uniforme e bem distribuída ao longo do ano e com temperaturas do mês mais frio entre 3 e 18ºC. Os dados climáticos registrados durante o período de avaliação das plantas (11-06 a 05-08-2003) na Estação Experimental da Cascata da Embrapa Clima Temperado foram: precipitação acumulada de 207,8 mm, temperatura média de 12,9 ºC, umidade relativa média de 71 % e 129 horas de frio acumulado (<7,2ºC).
Os ramos foram coletados em quatro datas (11-06, 11-07, 29-07 e 05-08), durante o período de dormência das plantas, no inverno de 2003, sendo transportados em caixa de isopor com gelo até o Laboratório de Fisiologia Vegetal da Embrapa Clima Temperado, onde os tecidos foram separados, congelados em nitrogênio líquido, armazenados em ultrafreezer (-70ºC), liofilizados e moídos para serem analisados.
A metodologia para a análise das concentrações dos açúcares solúveis (sacarose, sorbitol, glicose e frutose), em mg g-1 MS liofilizada, foi padronizada para tecidos vegetais de frutíferas no Laboratório de Fisiologia Vegetal da Embrapa Clima Temperado, a partir da adaptação da metodologia adotada por Rakngan (1995). Para a extração dos açúcares solúveis, amostras secas e moídas de 500 mg foram incubadas por 5 min a 80-85ºC, em 5 ml de etanol, a 80% e centrifugadas a 6.000 rpm, por 10 min. O sobrenadante foi coletado e o precipitado novamente ressuspenso e centrifugado em 10 ml de etanol 80%. O segundo sobrenadante foi coletado e misturado ao primeiro, enquanto o precipitado foi armazenado para a análise da concentração de amido. A mistura dos sobrenadantes foi evaporada em rotavapor a 45ºC, até não conter mais álcool. As amostras foram diluídas em água deionizada e filtradas em colunas contendo resina IR 120 (catiônica), para eliminar aminoácidos e, a seguir, em resina IR 400 (aniônica) para eliminar ácidos orgânicos. Após eluição das amostras, as mesmas foram decantadas para a precipitação e remoção das proteínas e filtradas em papel-filtro qualitativo. O filtrado foi evaporado em cadinho de porcelana mantido a 240-270ºC, até secar. Em seguida, adicionou-se 1ml de pentaerythritol (1mg.ml-1, açúcar-padrão interno) e, novamente, submeteu-se ao aquecimento (200ºC), até a completa secagem. Adicionaram-se 1ml de pyridine, para solubilização, 200µL de hexamethyldisilasane (HMDS) e 100 µL de trimethylchlorosilane (TMCS), para a metilação, até que ocorresse a formação de um precipitado branco. Após o resfriamento, 2 µL da amostra foram injetados no aparelho. Os padrões utilizados e respectivos tempos de retenção foram: padrão interno de pentaerythritol 5'48", frutose 10'95", a-glicose 12'49", sorbitol 13'50", b-glicose 14'11" e sacarose 29'94".
A concentração de amido, em mg g-1 MS liofilizada, foi obtida de acordo com a metodologia de McCready et al. (1950) por espectrofotometria. Do precipitado obtido na extração dos açúcares solúveis, 100 mg de peso seco foram ressuspensos em ácido perclórico a 30% e centrifugados a 3.000 rpm, durante 10 minutos. O mesmo procedimento foi repetido por mais uma vez. Os sobrenadantes foram coletados e misturados, e o precipitado, descartado. As amostras foram misturadas ao reativo de antrona, fervidas durante 10 min e resfriadas. A leitura da absorbância foi realizada em 620 nm.
O delineamento experimental foi inteiramente casualizado, com três repetições, em esquema fatorial AxB, sendo o fator A: data de coleta (11-06, 11-07, 29-07 e 05-08) e o fator B: tipo de tecido (gema e base da gema). As variáveis analisadas nos experimentos foram as seguintes concentrações: amido, sacarose, sorbitol, glicose, frutose e açúcares solúveis totais. As médias foram comparadas pelo teste de Duncan, a 5% de probabilidade, empregando-se o programa estatístico SANEST (Zonta & Machado, 1991).
RESULTADOS E DISCUSSÃO
Experimento 1 Plantas sem sintomas de morte-precoce
Ocorreu interação significativa (P<0,05) entre os fatores para as concentrações de amido (%), açúcares solúveis totais, glicose e frutose (mg g-1 MS liofilizada) e não ocorreu interação para as concentrações de sacarose e sorbitol nas plantas sem sintomas de morte-precoce. A concentração de amido (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, durante as três primeiras coletas (11-06, 11-07 e 29-07) e nas gemas, durante a última coleta (05-08). A concentração de amido aumentou nas gemas, nas duas últimas coletas (29-07 e 05-08) e na base das gemas, na segunda coleta (11-07) (Figura 1a).

A elevada concentração de amido na base das gemas durante a dormência, e o aumento na concentração de amido nas gemas, na fase da saída de dormência, propiciaram um adequado suprimento energético dos tecidos para que a brotação e a floração destas plantas ocorressem de maneira uniforme e regular. O amido é o principal carboidrato de reserva das plantas frutíferas, sendo inicialmente consumido por conversão, em açúcares solúveis, os quais são usados no processo de respiração para suprimento de energia para brotação e floração (Stassen, 1980).
Marquat et al. (1999) afirmaram que, durante a dormência, a gema possui baixa capacidade mobilizadora para assimilados e nutrientes minerais, aumentando a hidrólise do amido e a síntese de sacarose como forma de proteção ao frio e que, no período de saída da dormência, a gema se torna um dreno metabólico preferencial, acumulando reservas (sorbitol e amido) por meio de transporte ativo, as quais seriam metabolizadas, induzindo o rompimento da gema.
A concentração de açúcares solúveis totais (mg g-1 MS liofilizada) foi superior na base das gemas, nas duas primeiras coletas (11-06 e 11-07). A concentração de açúcares solúveis totais aumentou em ambos os tecidos, na segunda coleta (11-07) e diminuiu na base das gemas, na terceira coleta (29-07) (Figura 1b).
A concentração de açúcares solúveis totais aumentou nas gemas e diminuiu na base das gemas, na metade da fase de dormência, possivelmente, para o suprimento energético dos tecidos na fase da retomada de crescimento das gemas. Flore & Laine (1996) afirmaram que, no pessegueiro, os carboidratos totais são armazenados principalmente em ramos, atingindo um máximo na metade do período de repouso.
A concentração de glicose (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, na primeira coleta (11-06) e nas gemas, na terceira coleta (29-07). A concentração diminuiu na base das gemas, na segunda coleta (11-07) sendo que, nas gemas, houve aumento na terceira coleta (29-07) e diminuição na última (05-08) (Figura 1c).
A concentração de frutose (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, nas duas primeiras coletas (11-06 e 11-07) e nas gemas, na terceira coleta (29-07). A concentração aumentou nas gemas e diminuiu na base das gemas na terceira coleta (29-07) e sofreu redução nas gemas, na última coleta (05-08) (Figura 1d).
As concentrações de glicose e frutose diminuíram na base das gemas e aumentaram nas gemas, na metade do período de repouso, sendo que, na fase da saída de dormência, a mobilização destes açúcares aumentou nas gemas, possivelmente, para a obtenção de energia e o fornecimento de esqueletos carbônicos para a retomada do crescimento ativo. Sturm (1999) demonstrou que são necessários altos níveis de hexoses para a manutenção da pressão osmótica da célula na fase do elongamento celular.
Experimento 2 Plantas com sintomas de morte-precoce
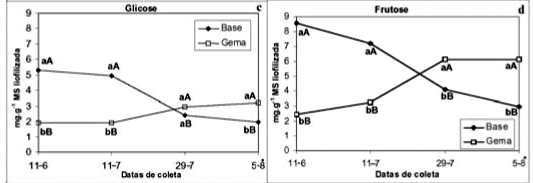
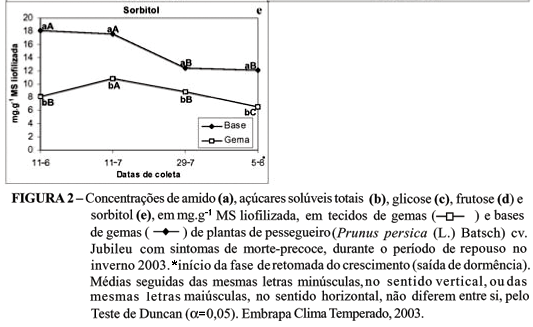
Ocorreu interação significativa (P<0,05) entre fatores para as concentrações de amido (%), açúcares solúveis totais, glicose, frutose e sorbitol (mg g-1 MS liofilizada) e não ocorreu interação para a concentração de sacarose nas plantas com sintomas de morte-precoce.
A concentração de amido (%) foi significativamente superior na base das gemas nas duas primeiras coletas (11-06 e 11-07) A concentração de amido aumentou nas gemas, na terceira coleta (29-07) e diminuiu em ambos os tecidos, na última coleta (05-08) (Figura 2a).

A antecipação na quebra de dormência provocada pelo desencadeamento da síndrome da morte-precoce, nestas plantas, intensificou a degradação do amido em ambos os tecidos, na fase da saída de dormência, possivelmente, para o fornecimento de glicose aos tecidos na fase da retomada do crescimento ativo das gemas. A degradação do amido pode estar relacionada com o fornecimento de glicose aos tecidos para a manutenção de um gradiente de concentração e a proteção em períodos de frio excessivo (Preiss & Sivak, 1996).
A concentração de açúcares solúveis totais (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, durante as três primeiras coletas (11-06, 11-07 e 29-07). A concentração aumentou nas gemas, na segunda coleta (11-07) e diminuiu na base das gemas, na terceira coleta (29-07) (Figura 2b).
A concentração de açúcares solúveis totais aumentou nas gemas, no início, e diminuiu na base das gemas, na metade do período de repouso, possivelmente, devido à degradação da sacarose e do sorbitol para o fornecimento de glicose e frutose aos tecidos. De acordo com os resultados encontrados por Rakngan (1995) em gemas de pereira cultivar Nijisseiki, a concentração de açúcares solúveis aumenta quando as plantas entram em repouso e diminuem antes ou durante a brotação.
A concentração de glicose (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, nas duas primeiras coletas (11-06 e 11-07) e nas gemas, na última coleta (05-08). A concentração de glicose aumentou nas gemas e diminuiu na base das gemas, na terceira coleta (29-07) (Figura 2c).
A concentração de frutose (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, nas duas primeiras coletas (11-06 e 11-07) e nas gemas, nas duas últimas coletas (29-07 e 05-08). A concentração de frutose diminuiu na base das gemas e aumentou nas gemas, na terceira coleta (29-07) (Figura 2d).
As concentrações de glicose e frutose diminuíram na base das gemas e aumentaram nas gemas a partir da metade do período de repouso, sendo que estas concentrações se mantiveram elevadas nas gemas até a fase de saída da dormência. Yamaki & Ishikawa (1986) verificaram que frutose e glicose são principalmente produzidas a partir da degradação do sorbitol em tecidos de macieira.
A concentração de sorbitol (mg g-1 MS liofilizada) foi significativamente superior na base das gemas, em todas as coletas (11-06, 11-07, 29-07 e 05-08). A concentração nas gemas aumentou na segunda coleta (11-07) e diminuiu nas duas últimas coletas (29-07 e 05-08). Na base das gemas, a concentração sofreu redução na terceira coleta (29-07) (Figura 2e).
A mobilização do sorbitol, em ambos os tecidos a partir da metade do período de repouso, estaria aumentando as concentrações de glicose e frutose nas gemas para o suprimento energético das gemas na fase de saída da dormência . O sorbitol é o açúcar dominante na seiva do floema das plantas da família Rosaceae, sendo sua principal função proteger os tecidos contra a desidratação, a salinidade e o frio, caracterizando-se também como um carboidrato de reserva (Grant & Rees, 1981).
CONCLUSÕES
1. A base das gemas demonstrou ser um importante tecido de reserva de carboidratos para a retomada do crescimento das gemas no início da primavera, em ambos os pomares.
2. A elevada concentração de amido nos tecidos das plantas sem sintomas da morte-precoce, na fase da saída de dormência, propiciou um adequado suprimento energético para que a brotação e a floração destas plantas ocorressem de maneira uniforme e regular.
3. A antecipação na quebra de dormência das plantas com sintomas de morte-precoce intensificou a degradação de amido e sorbitol nos tecidos, na saída de dormência, para o fornecimento de glicose e frutose, que estariam sendo metabolizadas para o suprimento energético na fase de rompimento das gemas e a proteção dos tecidos contra o resfriamento.
Recebido em 21-07-2006. Aceito para publicação em 25-01-2007.
- CAMPOS, A.D.; CARVALHO, F.L.C. Efeito da falta e do excesso de água, associado a variações bruscas de temperatura na morte precoce de pessegueiros. In: CPNGRESSO BRASILEIRO DE FRUTICULTURA, 13., Anais... Salvador: SBF, 1994.
- FLORE, J.A.; LAINE, D. R. Prunus. In: ZAMSKI, E.; SCHAFFER, A.A. Photoassimilate distribution in plants and crops: source-sink relationships. New York: Marcel Dekker, 1996, p. 825 - 849.
- FORTES, J.F.; OSÓRIO, V.A. Pêssego: fitossanidadae. Pelotas-RS. Embrapa Clima Temperado, Brasília: Embrapa Informação Tecnológica, 2003. 53 p.
- GRANT, C.R.; REES, T. Sorbitol metabolism by apple seedlings. Phytochemistry, New York, v. 20, p. 1505-1511, 1981.
- MARQUAT, C.; VANDAMME, M.; GENDRAUD, M.; PÉTEL, G. Dormancy in vegetative buds of peach: relation between carbohydrate absortion potentials and carbohydrate concentration in the bud during dormancy and its release. Scientia Horticulturae, Hamsterdan v. 79, p. 151-162, 1999.
- McCREADY, R.M.; GUGGOLZ, J.; SILVEIRA, V. OWENS, H.H. Determination of starch and amylose in vegetables. Analytical Chemistry, Washington, v. 22: 1156-1158, 1950.
- MEDEIROS, C.A.B.; RASEIRA, M.C. A Cultura do pessegueiro Embrapa Clima Temperado Pelotas-RS, p. 280 296,1998.
- PREISS, J.; SIVAK, M. N.; Starch synthesis in sink and sources. In: ZAMSKI, E.; SCHAFFER, A.A. Photoassimilate distribution in plants and crops: source-sink relationships. New York: Marcel Dekker, 1996, p. 63-93.
- RAKNGAN, J. Carbohydrate analysis of Japanese pear trees grown under adverse conditions. Phenological and physiological study of Japanese pear grown under adverse condition, Tsukuba, 1995, cap. 3, p. 61-63.
- STASSEN, P.J.C. Reserves in deciduous plant trees and implications in deciduous fruit grower. Deciduous Plant Grower, Cape Town, v. 30, p. 467-472, 1980.
- STURM, A. Invertases. Primary Structures, Functions, and Roles in Plant Development and Sucrose partitioning Plant Physiology, Califórnia, v. 121, p. 1-7, 1999.
- YAMAKI, S.; ISHIKAWA, K. Roles of four sorbitol related enzymes and invertase in the seasonal alteration of sugar metabolism in apple tissue The Journal of American Society for Horticultural Sciences, Mount Vernon, v.111, p.134-137, 1986.
- ZONTA, E.P.; MACHADO, A.A. Manual do SANEST: sistema de análise estatística para microcomputadores. Pelotas: UFPel, 1991. 102p.
Datas de Publicação
-
Publicação nesta coleção
26 Jun 2007 -
Data do Fascículo
Abr 2007
Histórico
-
Aceito
25 Jan 2007 -
Recebido
21 Jul 2006